Le tadalafil possède une affinité marquée pour la PDE5, mais épargne en grande partie les isoformes PDE1, PDE2 et PDE11, réduisant ainsi le risque d’effets extra-caverneux. L’action se traduit par une augmentation contrôlée de la circulation sanguine locale, indépendante des variations alimentaires. Sa pharmacocinétique repose sur une absorption digestive rapide, un métabolisme hépatique par CYP3A4 et une distribution tissulaire large. La biodisponibilité reste stable, et l’équilibre plasmatique est atteint en quelques jours lors d’administrations répétées. Les interactions cliniquement significatives surviennent avec les inhibiteurs puissants de CYP3A4 tels que le kétoconazole. Dans la littérature pharmacologique, acheter cialis 20 mg est souvent associé à des schémas d’utilisation basés sur la durée prolongée de son action.
Oldweb.northampton.ac.uk
FORUM is intended for new ideas or new ways of interpreting existing information. It
provides a chance for suggesting hypotheses and for challenging current thinking onecological issues. A lighter prose, designed to attract readers, will be permitted. Formal
research reports, albeit short, will not be accepted, and all contributions should be concise
with a relatively short list of references. A summary is not required. Latitudinal trends in plant-pollinator interactions: are tropical plants more specialised? Jeff Ollerton and Louise Cranmer, School of En7ironmental Science, Uni7. College Northampton, Park Campus, Northampton, NN2 7AL, UK ( jeff.ollerton@northampton.ac.uk). The increase in richness of species and higher taxa going from
diversity and consequent finer division of resources, polar
higher to lower latitudes is one of the most studied global
areas because of low species diversity and therefore a lack
biogeographical patterns. Latitudinal trends in the interactions
of opportunity for species to be more generalised. Are
between species have, in contrast, hardly been studied at all, probably because recording interactions is much less straightfor-
there any data for global patterns of species interactions
ward than counting species. We have assembled two independent data sets which suggest that plant-pollinator interactions are not
Few quantitative studies have explicitly addressed the
more ecologically specialised in the tropics compared to temper-
question of how the specificity of species interactions
ate latitudes. This is in contrast to a prevailing view that tropical ecological interactions tend towards higher specificity than their
varies with latitude. For example, Scriber (1973) and
temperate counterparts.
Price (1980) looked at specialisation in larval feeding inLepidoptera, Beaver (1978) dealt with bark and ambrosiabeetles, Hawkins (1990) and Porter and Hawkins (1998)
Latitudinal trends in biodiversity are well known for
studied global patterns of parasitoid numbers per insect
many groups of organisms, with taxon diversity being
host, whilst Rohde (1978) focused on latitudinal trends
in fish parasites. Their findings will be considered later,
(MacArthur 1972, Rohde 1992, Gaston and Williams
but the question of tropical ecological specialisation
1996). In contrast, we know much less about latitudinal
remains largely unresolved for most categories of inter-
trends in the biodiversity of species interactions. For
action and the functional groups involved in these
example, as one moves from temperate to tropical
latitudes, do predator-prey, parasite-host or mutualistic
Despite their importance in most terrestrial ecosystems
(Kearns and Inouye 1997), we possess an inadequate
(defined as the number of species involved in the interac-
knowledge of the broad biogeographic patterns of plant-
tion, as distinct from morphological or evolutionary
pollinator interactions and the underlying causes of any
specialisation; Waser et al. 1996, Armbruster et al. 2000)?
pattern (Johnson and Steiner 2000). An initial reading of
The proposal that resources are divided more finely
the literature would suggest that there is a consensus
amongst a greater number of species in the tropics,
amongst pollination biologists that tropical pollination
compared to temperate communities (MacArthur 1972,
systems are more ecologically specialised than temperate
Janzen 1973), suggests that tropical organisms should
systems (Johnson and Steiner 2000) but there are almost
indeed be more ecologically specialised. However, low
no data to support this assertion, and only limited data
species diversity in very high latitude areas may also lead
to refute it (Kevan and Baker 1983).
to apparent ecological specialisation in species interac-tions. In this case, the resulting latitudinal trend wouldbe hump-backed – high specificity of interactions in the
The data sets
tropics and towards polar regions, with much lowerspecificity (greater generalisation) at temperate latitudes.
As a step towards understanding whether pollination
Interestingly, the extremes of the gradient would show
systems show a significant latitudinal trend in speciali-
greater specialisation in interactions for diametrically
sation, we have assembled two independent data sets at
opposite reasons – the tropics because of high species
different taxonomic/ecological scales, full details of
which are given in Appendices 1 and 2. The first data
variance in this regression model is explained by differ-
set is at the scale of the plant community and comprises
ences in sampling effort between communities. Clearly
27 published and unpublished surveys of plant-flower
tropical community pollination studies suffer from un-
visitor interactions in 35 communities at different lati-
der sampling of the true diversity of flower visitors per
tudes. From these studies we extracted information on
plant species (though note that latitude and sampling
the latitude at which the study was undertaken (deci-
effort are not directly correlated – Pearson’s Product
malised for the purposes of analysis), mean number of
Moment Correlation: r = 0.05, df = 33, p = 0.78).
species of flower visitors per plant species (most of these
The distribution of the asclepiad data set is highly
studies recorded flower visitors rather than pollinators
non-normal and untransformable and therefore violates
per se; however, number of flower visitors is strongly
the requirements of multiple regression analysis. To
correlated with number of pollinators and this should
take account of sampling effort for these data we have
therefore be an appropriate proxy [Ollerton, unpubl.]),
corrected number of pollinators per plant species by
number of plant species studied and sampling effort
dividing by number of days sampling for the subset of
(number of field days of observation). The latter vari-
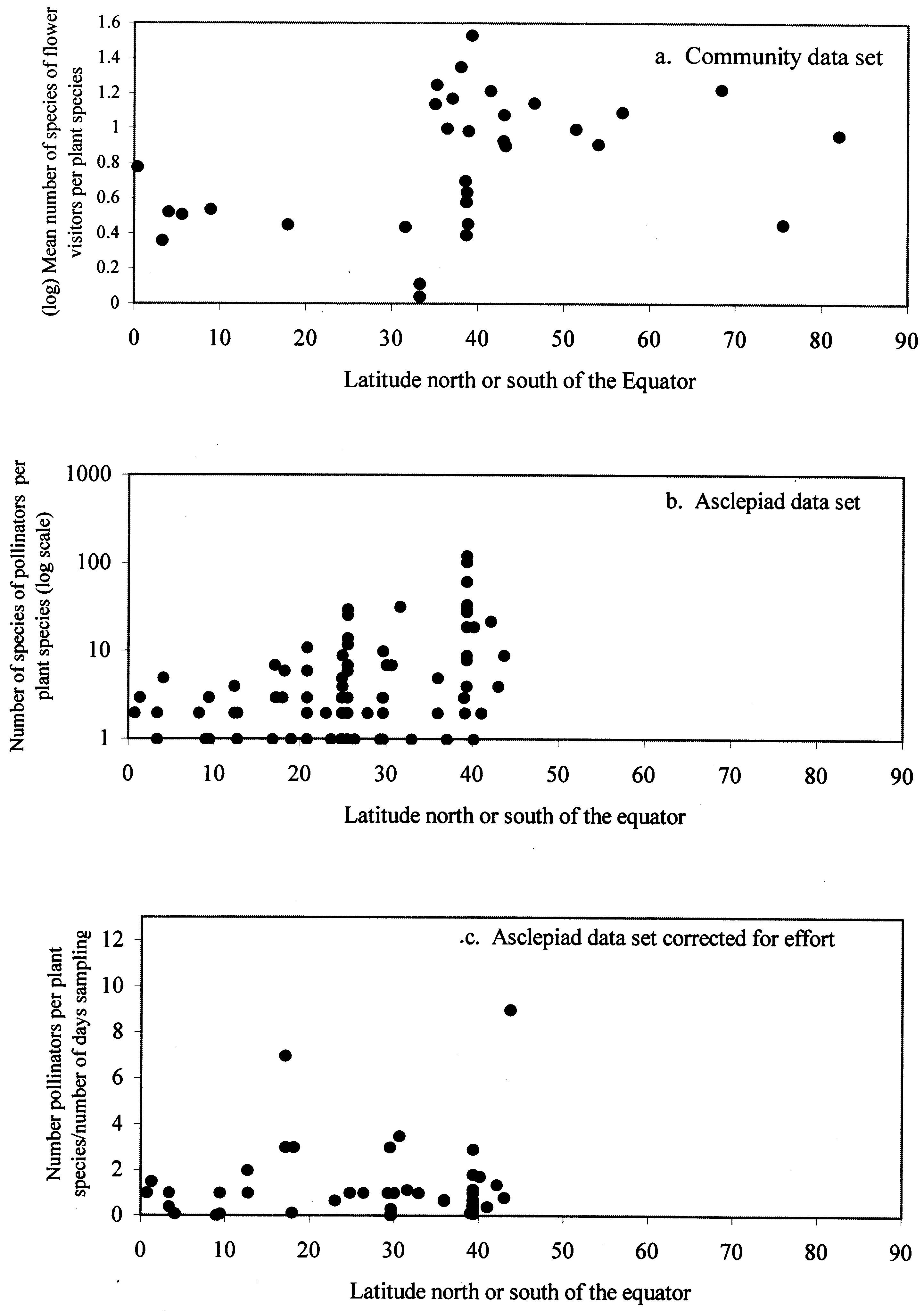
data where this is known (Fig. 1c). Correcting for
able was in some studies explicitly stated and in others
sampling effort in this way removes any correlation
was estimated from the published information.
between latitude and pollinator specialisation. Once
The second data set consists of 103 published and
again, the apparently more specialised tropical species
unpublished studies of pollinators of species of asclepi-
suffer from under sampling of pollinators.
ads (subfamily Asclepiadoideae of the Apocynaceae
Two completely independent data sets, at two differ-
sensu Endress and Bruyns 2000). This is part of the
ent taxonomic scales, show precisely the same result,
that tropical plants are, on average, no more ecologi-
bayreuth.de/departments/planta2/wgl/fsigrid2.html). As
cally specialised in their pollination systems than tem-
in the first data set, we extracted information on lati-
perate species. We conclude that the apparent trend
tude, number of pollinators per plant species (in this
towards more specialised pollination systems for tropi-
case, a much more straightforward variable as asclepi-
cal plants shown in Fig. 1a and b is an artefact of
ads possess aggregations of pollen (pollinia) that me-
sampling bias and that there is no significant latitudinal
trend in the specificity of plant-pollinator relationships.
identification of pollinators much easier – see Ollerton
How do our results compare to the previously pub-
and Liede 1997) and number of days of observation,
lished studies cited earlier. In particular, is there any
which was available for only 59 of the 103 studies.
evidence from other work that the humpbacked latitu-dinal trend may occur in some interactions? Thesestudies have looked at a range of organisms and typesof interaction and have uncovered a variety of relation-
Results and discussion
ships between latitude and ecological specificity. Scriber(1973) was probably the first worker to confront quan-
Initial analyses of these data sets suggests that pollina-
titatively the problem of temperate versus tropical spe-
tion systems do indeed become more specialised moving
cialisation, in a study of larval host plant use in
from temperate latitudes towards the tropics (Fig. 1a
Papilionidae (Lepidoptera). His analysis showed that a
and b). In both the community and asclepiad data sets
higher proportion of temperate species could be consid-
there is a significant positive relationship between lati-
ered generalist compared to tropical species. Scriber’s
tude and number of pollinators/flower visitors per plant
definition of generalist taxa was ‘‘…those species feed-
species. This is also true if the data are separated into
ing on more than one taxonomic family of plants…’’.
northern and southern hemispheres (data not pre-
This may be considered a rather broad definition of
sented). However, closer analysis reveals that this pat-
‘‘generalised’’ and, intriguingly, Price (1980) presented
tern is misleading. The various studies included within
data that suggested that tropical butterflies tended to be
the community and asclepiad data sets varied consider-
no more host specific than temperate species. Rohde
ably in the sampling effort undertaken to observe and
(1978) found that tropical taxa of marine platyhelminth
record flower visitors. To take account of this we have
fish parasites in the group Digenea were more host
used sampling effort per plant species together with
specific than temperate taxa, but that this was not so in
latitude as independent variables in a multiple regres-
the Monogenea. Beaver (1978) showed that bark and
sion analysis of the community data set (Table 1).
ambrosia beetles (Coleoptera: Scolytidae and Platypo-
Forty percent of the variation in mean number of
didae) are actually less host specific in the tropics
flower visitors per plant species is explained by this
compared to temperate communities, a pattern that he
stepwise multiple regression model. However, only 4%
considered may be explained by the low population
of this variation results from the latitude at which the
densities of host trees in the tropics. Hawkins (1990)
study was conducted (and this is only significant at
studied parasitoids of phytophagous insects with differ-
p = 0.075 for the t-ratio test). The remaining 36% of the
ent feeding ecologies and showed that those parasitising
exposed hosts tended to be more host specific in the
same’’. This conclusion is confirmed by the data that we
tropics, whilst no such pattern was apparent for para-
have presented in this paper. Tropical communities
sitoids utilising hosts concealed in plant tissue. Clearly,
provide some of the best examples of close co-evolved
different categories of species interaction and different
plant-pollinator relationships and in absolute terms do
groups of taxa may or may not show increased special-
contain a higher number of plants with specialised
pollination systems. However, tropical plant assem-
A literature review by Kevan and Baker (1983) con-
blages are on average many times more species-rich than
cluded that ‘‘…from the arctic and alpine areas to the
their temperate counterparts and so may not in fact
lowland tropics, it appears that the frequency of occur-
possess disproportionately more ecologically specialised
rence of specialised pollination syndromes is about the
pollination systems than temperate assemblages.
Fig. 1. Relationshipsbetween latitude andpollinator specialisationfor the communitysurvey and asclepiaddata sets. a. Community surveysof plant-flower visitorrelationships. Meannumber of species offlower visitors per plantspecies has been logtransformed. Pearson’sproduct momentcorrelation: r = 0.33,df = 33, p = 0.051. b. Pollinators ofasclepiads. Spearmanrank correlation:r = 0.33, n = 91,p = 0.002. c. Pollinatorsof asclepiads, correctedfor sampling effort. Spearman rankcorrelation: r = 0.09,n = 59, p = 0.51.
Table 1. Results of stepwise multiple regression on mean number of species of flower visitors per plant species for thecommunity data set. All variables were natural log transformed. Problems with the data sets
plant species, localities and years and so there is nosimple ‘‘rule of thumb’’ which would allow us to apply
The type of analysis that we have presented, in which
a simple correction. We have therefore opted to use a
largely pre-existing data are evaluated in relation to a
range of days of sampling effort to test how a saturat-
question which they were not primarily collected to
ing sampling function would affect our conclusions. We
address, can be fraught with statistical problems. We
repeated the analysis of the asclepiad data set using
have identified two possible causes for concern within
sampling saturation points between 1 day and 60 days
the two data sets, which we detail below.
of sampling effort (Table 2). This covered the range of
The first statistical problem concerns the phyloge-
numbers of days of actual sampling effort undertaken
netic relatedness of the plants and pollinators in the
by the various studies in Appendix 2. The analysis
analysis. It is acknowledged (and debated) that possible
involved repeating the Spearman rank correlations be-
phylogenetic biases must be taken into consideration in
tween number of species of pollinator (corrected for
any comparative analysis (Harvey and Pagel 1991).
sampling effort) and latitude and successively restricting
However, the community survey data set spans such a
the maximum number of days by which number of
wide range of plant and animal genera, orders and
pollinators was corrected to 1, 2 … 10 … 20 … up to
classes that a formal phylogenetically-corrected regres-sion is not possible. Whether it is required for such a
60 days. Low levels of maximum sampling effort (less
phylogenetically broad spread of taxa is arguable. In
than 10 days) yielded results not quantitatively different
relation to the asclepiad data set, a robust molecular
from that shown in Fig. 1b, with statistically significant
generic-level phylogeny of the group is not yet avail-
relationships between latitude and number of species of
able. Therefore, whilst we recognise that the phyloge-
pollinator. That is to say, correcting by a maximum of
netic architecture of this data set may be a statistical
only a modest sampling effort is approximately similar
problem (for example, the higher latitude data mainly
to not correcting the data at all, a not unexpected
come from North American Asclepias species) we can-
result. The statistically significant correlation disap-
not at the present time allow for this.
pears when using more realistic saturation levels of
The second statistical problem specifically concerns
the asclepiad data set. In order to correct for different
Table 2. Spearman rank correlations of latitude versus num-ber of species of pollinators per plant species corrected by
sampling efforts across studies, for each plant the num-
sampling effort for a range of sampling effort saturation
ber of recorded pollinators was divided by the number
points. N = 59 in all cases, except the uncorrected analysis,
of days of sampling. This correction assumes a linear
relationship between sampling effort and number of
pollinators per plant species. In reality the relationshipis likely to be saturating, with records of new pollina-
tors declining to zero at some point during the observa-
tion period. If the relationship between sampling effort
and number of observed pollinators is indeed saturat-
ing, our simple correction would result in an under
estimate of the number of pollinators per plant species
expected from a given level of sampling effort. It is
impossible to say what the exact sampling saturation
point is as this information is never presented in studies
of plant-pollinator interactions. In a recent survey of
asclepiad pollinators at a site in South Africa, we had
sampled all of the pollinators of some species in as little
as 10 days, though for other species we were still
recording new pollinators after 30 days (Ollerton et al.
in prep.). Sampling saturation points (beyond which no
new pollinators are recorded) are likely to vary between
exploring latitudinal trends in plant-pollinator interac-tions, these data sets are as good as any that couldbe currently assembled. We hope that by publishingthis study we will stimulate interest in the question oftropical versus temperate specialisation in ecologicalinteractions and that future researchers will obtaingrants large enough to allow dedicated data collectionthat will tackle this question. Until such time, thesedata sets must suffice. Acknowledgements – The ideas presented in this paper havebenefited from discussion with many colleagues. We wouldparticularly like to thank Scott Armbruster, Kevin Gaston,David Inouye, Steve Johnson, Duncan McCollin, Jane Mem-mott, Paul Neal, Jens Olesen and Nick Waser and an anony-mous reviewer. We also thank Steve Johnson, Sigrid Liede,Jane Memmott, Jens Olesen, Anton Pauw and Milene Vieirafor providing us with unpublished data. We are grateful to thefollowing organisations for providing funding which con-tributed to some the results in the paper: The Royal Society,The Leverhulme Trust, Church and Co. PLC, The BiodiversityTrust, The Percy Sladen Memorial Fund and The RoyalEntomological Society.
Fig. 2. The relationship between number of days sampling
References
effort and the number of flower visitors/pollinators per plantspecies in (a) the community data set; and (b) the asclepiad
Armbruster, W. S., Fenster, C. B. and Dudash, M. R. 2000.
Pollination ‘‘principles’’ revisited: specialization, pollina-tion syndromes and the evolution of flowers. – Det NorskeVidenskaps-Akademi I. Mat-Naturv. Klasse Skrifter Ny.
greater than 10 days sampling (Table 2) and confirm
Beaver, R. A. 1978. Host specificity of temperate and tropical
In conclusion, correcting the data for sampling ef-
animals. – Nature 281: 1139 – 1141.
Endress, M. E. and Bruyns, P. V. 2000. A revised classification
fort using a realistic saturating function (by which we
of the Apocynaceae s.l. – Bot. Rev. 66: 1 – 56.
consider that 10 days or less of observation is un-
Fishbein, M. and Venable, D. L. 1996. Diversity and temporal
likely to identify all of the pollinators of even a mod-
change in the effective pollinators of Asclepias tuberosa. –Ecology 77: 1061 – 1073.
erately generalised species) does not affect the results
Gaston, K. J. and Williams, P. H. 1996. Spatial patterns in
obtained when a linear, non-saturating correction is
taxonomic diversity. – In: Gaston, K. J. (ed.), Biodiversity:
applied. This raises quite a fundamental issue in rela-
a biology of numbers and difference. Blackwell Scientific,pp. 202 – 229.
tion to studying pollination ecology – when can we
Hawkins, B. A. 1990. Global patterns of parasitoid assemblage
be sure that we have identified all of the pollinators
size. – J. Anim. Ecol. 59: 57 – 72.
of a plant? The annual fluctuations in pollinator
Harvey, P. H. and Pagel, M. 1991. The comparative method in
abundances that are a feature of many plant-pollina-
evolutionary biology. – Oxford Univ. Press.
Janzen, D. H. 1973. Comments on host-specificity of tropical
tor systems (see, amongst many potential examples,
herbivores and its relevance to species richness. – In:
Pettersson 1991, Fishbein and Venable 1996, Lam-
Heywood, V. H. (ed.), Taxonomy and ecology. Academic
born and Ollerton 2000) suggests that a time scale of
Johnson, S. D. and Steiner, K. E. 2000. Generalization versus
years to decades may be necessary before a complete
specialization in plant pollination systems. – Trends Ecol.
list of pollinators is obtained for generalist pollination
systems. This is reinforced by a crude analysis com-
Kearns, C. A. and Inouye, D. W. 1997. Pollinators, flowering
plants, and conservation biology. – BioScience 47: 297 –
paring sampling effort to number of identified polli-
nators in the community and asclepiad data sets
Kevan, P. G. and Baker, H. G. 1983. Insects as flower visitors
presented here (Fig. 2a and b). In both of the data
and pollinators. – Ann. Rev. Ent. 28: 407 – 453.
Lamborn, E. and Ollerton, J. 2000. Experimental assessment
sets there is no suggestion of a levelling off of num-
of the functional morphology of inflorescences of Daucus
bers of identified pollinators as sampling effort in-
carota (Apiaceae): testing the ‘‘fly catcher effect’’. – Funct.
We have attempted to be honest about the limita-
MacArthur, R. H. 1972. Geographical ecology. – Harper and
tions of our data sets and would argue that apprecia-
Ollerton, J. and Liede, S. 1997. Pollination systems in the
tion of these problems does not negate their value,
Asclepiadaceae: a survey and preliminary analysis. – Biol.
nor the value of our analyses. For the purposes of
Pettersson, M. W. 1991. Pollination by a guild of fluctuating
marine Monogenea and Digenea. – Mar. Biol. 47: 125 – 134.
moth populations: option for unspecialization in Silene
Rohde, K. 1992. Latitudinal gradients in species diversity: the
6ulgaris. – J. Ecol. 79: 591–604.
search for the primary cause. – Oikos 65: 514 – 527.
Porter, E. E. and Hawkins, B. A. 1998. Patterns of diversity for
Scriber, J. M. 1973. Latitudinal gradients in larval feeding
aphidiine (Hymenoptera: Braconidae) parasitoid assem-
specialization of the world Papilionidae (Lepidoptera). –
blages on aphids (Homoptera). – Oecologia 116: 234 – 242.
Price, P. W. 1980. Evolutionary biology of parasites. – Prince-
Waser, N. M., Chittka, L., Price, M. V. et al. 1996. Generaliza-
tion in pollination systems, and why it matters. – Ecology
Rohde, K. 1978. Latitudinal differences in host-specificity of
Appendix 1. Studies included in the community-level data set.
Appendix 2. Studies used in the asclepiad data set. Note that data for sampling effort are not available for all studies. Ceropegia lushi var. acuminataOxypetalum alpinum var. alpinumOxypetalum banksii subsp. banksii
Ali, T. 1994. Pollination ecology of some asclepiads (Asclepiadaceae) from Pakistan. – Unpublished Ph.D. thesis, Univ. of Karachi. Arroyo, M. T. K., Primack, R., Armesto, J. 1982. Community studies in pollination ecology in the high temperate Andes of Chile. I. Pollination mechanisms and altitudinal variation. – Am. J. Bot. 69: 82 – 97. Barrett, S. C. H. and Helenurm, K. 1987. The reproductive biology of boreal forest herbs, I. Breeding systems and pollination. – Can. J. Bot. 65: 2036 – 2046. Bhatnagar, S. 1986. On insect adaptations for pollination in some asclepiads of Central India. – In: Kapil, R. P. (ed.) Pollination biology – an analysis. Inter-India Publications,New Delhi, pp. 37 – 57. Betz, R. F., Struven, R. D., Wall, J. E. and Heitler, F. B. 1994. Insect pollinators of 12 milkweed (Asclepias) species. – In: Wickett, R. G., Lewis, P. D., Woodliffe, A. and Pratt,P. (eds.) Proc. Thirteenth N. Am. Prairie Conf. Dept of Parks and Recreation, Ontario, Canada, pp. 45 – 60. Bosch, J., Retana, J. and Cerda´, X. 1997. Flowering phenology, floral traits and pollinator composition in a herbaceous Mediterranean plant community. – Oecologia 109: 583 – 591. Burkill, I. H. 1897. Fertilization of some spring flowers on the Yorkshire coast. – J. Bot. 35: 92 – 189. Chaplin, S. J. and Walker, J. L. 1982. Energetic constraints and adaptive significance of the floral display of a forest milkweed. – Ecology 63: 1857 – 1870. Chaturvedi, S. K. and Pant, D. D. 1986. Further studies in the pollination of some Indian asclepiads. – Bull. Bot. Survey India 28: 23 – 30. Clements, F. E. and Long, F. L. 1923. Experimental pollination – an outline of the ecology of flowers and insects. – Carnegie Institution. Drapalik, D. J. 1969. A biosystematic study of the genus Matelea in the southeastern United States. – Unpublished Ph.D. thesis, Univ of North Carolina. Eberling, H. and Olesen, J. M. 1999. The structure of a high latitude plant-flower visitor system: the dominance of flies. – Ecography 22: 314 – 323. Fishbein, M. and Venable, D. L. 1996. Diversity and temporal change in the effective pollinators of Asclepias tuberosa. – Ecology 77: 1061 – 1073. Forster, P. I. 1989. Pollination of Marsdenia fraseri (Asclepiadaceae) by Metriorrhynchus lateralis (Coleoptera: Lycidae). – Coleopt. Bull. 43: 311 – 312. Forster, P. I. 1991. A possible identification for ‘‘Pollinia attached to adult anopheline mosquitoes from northern Australia’’. – Entom. Soc. Queensl. News Bull. 18: 113. Forster, P. I. 1992. Insects associated with the flowers of Marsdenia cymulosa Benth. (Asclepiadaceae) and their possible role in pollination. – Aust. Entom. Mag. 19: 45 – 47. Hagerup, O. 1932. On pollination in the extremely hot air at Timbuctoo. – Dansk. Bot. Ark.8: 1 – 20. Herrera, J. 1988. Pollination relationships in southern Spanish mediterranean shrublands. – J. Ecol. 76: 274 – 287. Hocking, B. 1968. Insect-flower associations in the high Arctic with special reference to nectar. – Oikos 19: 359 – 388. Inoue, T., Kato, M. and Kakutani, T. 1990. Insect-flower relationship in the temperate deciduous forest of Kibune, Kyoto: an overview of the flowering phenology and seasonalpattern of insect visits. – Contr. Biol. Lab. Kyoto Univ. 27: 377 – 463. Inouye, D. W. and Pike, G. H. 1988. Pollination biology in the snowy mountains of Australia: comparisons with montane Colorado, USA. – Aust. J. Ecol. 13: 191 – 210. Jonkers, B. 1990. Carallumas – gems of the mountainside. – Petr. Develop. Oman News 2: 7 – 11. Jonkers, B. 1993. De bestuivers van succulenten. – Succulenta 72: 268 – 275. Kanstrup, J. and Olesen, J.M. 2000. Plant-flower visitor interactions in a neotropical rain forest canopy: community structure and generalisation level. – Det NorskeVidenskaps-Akademi I. Matematisk-Naturvitenskapelig Klasse, Avhandlinger, Ny Serie 39: 33 – 41. Kato, M., Kakutani, T., Inoue, T. and Itino, T. 1990. Insect-flower relationship in the primary beech forest of Ashu, Kyoto: an overview of the flowering phenology and seasonalpattern of insect visits. – Contr. Biol. Lab. Kyoto Univ. 27: 309 – 375. Kevan, P. G. 1970. High arctic insect-flower relations: the inter-relationships of arthropods and flowers at Lake Hazen, Ellesmere Island, Northwest Territories, Canada. –Unpublished Ph.D. thesis, Univ. of Alberta, Canada. Kunze, H. and Liede, S. 1991. Observations on pollination in Sarcostemma (Asclepiadaceae). – Plant Syst. Evol. 178: 95 – 105. Liede, S. 1994. Some observations on pollination in Mexican Asclepiadaceae. – Madron˜o 41: 266 – 276. Liede, S. and Whitehead, V. 1991. Studies in the pollination biology of Sarcostemma 6iminale R.BR. sensu lato. – South Afr. J. Bot. 57: 115–122.
Lynch, S. P. 1977. The floral ecology of Asclepias solanoana woods. – Madrofio 24: 159 – 177. Macior, L. W. 1965. Insect adaptation and behaviour in Asclepias pollination. – Bull. Torrey Bot. Club 92: 114 – 126. Memmott, J. 1999. The structure of a plant-pollinator food web. – Ecol. Lett. 2: 276 – 280.
Meve, U. and Liede, S. 1994. Floral biology and pollination in stapeliads – new results and a literature review. – Plant Syst. Evol. 192: 99 – 116. Moldenke, A. R. and Lincoln, P. G. 1979. Pollination ecology in montane Colorado: a community analysis. – Phytologia 42: 349 – 379.
Momose, K., Yumoto, T., Nagamitsu, T. et al. 1998. Pollination biology in a lowland dipterocarp forest in Sarawak, Malaysia. I. Characteristics of the plant-pollinator communityin a lowland dipterocarp forest. – Am. J. Bot. 85: 1477 – 1501. Mosquin, T. and Martin, J. E. H. 1967. Observations on the pollination biology of plants on Melville Island, N.W.T., Canada. – Can. Field-Nat. 81: 201 – 205. Nel, M. 1995. Rare and interesting plants of the Namib desert, part 2: three desert plants. – Veld and Flora 81: 14 – 15. Pant, D. D., Nautiyal, D. D. and Chaturvedi, S. K. 1982. Pollination ecology of some Indian asclepiads. – Phytomorphology 32: 302 – 313. Pauw, A. 1998. Pollen transfer on birds’ tongues. – Nature 394: 731 – 732. Payson, E. 1916. The pollination of Asclepias cryptoceras. – Bot. Gaz. 61: 72 – 74. Percival, M. 1974. Floral ecology of coastal scrub in southeast Jamaica. – Biotropica 6: 104 – 129. Petanidou, T. 1991. Pollination ecology in a phryganic ecosystem. – Unpublished Ph.D. thesis, Univ of Thessaloniki. van der Pijl, L. 1954. Xylocopa and flowers in the tropics I, II and III. – Konink. Nederl. Akad. van Wetens, Proc. C. 57: 413 – 562. Piper, R. G., Sweeney, A. W. and Gibbons, D. S. 1990. Pollinia attached to adult anopheline mosquitoes from northern Australia. – Entom. Soc. Queensl. News Bull. 18: 83 – 84. Primack, R. B. 1983. Insect pollination in the New Zealand mountain flora. – New Zeal. J. Bot. 21:317 – 333Ramakrishna, T. M. and Arekal, G. D. 1979. Pollination biology of Calotropis gigantea (L.) R.Br. – Curr. Sci. 48: 212 – 213. Ramakrishna, T. M. and Arekal, G. D. 1982 – 83. Pollination in Pergularia daemia (Asclepiadaceae). – J. Mysore Univ. Sect. B. 29: 1 – 3. Rarnirez, N. 1989. Biologia de polinisacion en una comunidad arbustiva tropical de la Alta Guayana Venezolana. – Biotropica 21:319 – 330. Rarmirez, N. and Brito, Y. 1992. Pollination biology in a palm swamp community in the Venezuelan Central Plains. – Bot. J. Linn. Soc. 110: 277 – 302. Robertson, C. 1891. Flowers and insects, Asclepiadaceae to Scrophulariaceae. – Trans. St. Louis Acad. Sci. 5: 569 – 598. Robertson, C. 1928. Flowers and insects: lists of visitors of four hundred and fifty-three flowers. – Privately published, Carlinville, Illinois. Sabrosky, C. W. 1987. A new species of Leptometopa (Diptera, Milichiidae) from Madagascar pollinating Ceropegia (Asclepiadaceae). – Proc. Ent. Soc. Washington 89: 242 – 243. Skutch, A. F. 1988. Flowering and seed-production of Fischeria fimebris (Asclepiadaceae). – Brenesia 30: 13 – 17. Small, E. 1976. Insect pollinators of the Mer Bleue peat bog of Ottawa. – Can. Field-Nat. 90: 22 – 28. Struck, M. 1995. Land of blooming pebbles: flowers and their pollinators in the Knersvlakte. – Aloe 32: 56 – 64. Vieira, M. F. and Shepherd, G. F. 1999. Pollinators of Oxypetalum (Asclepiadaceae) in southeastern Brazil. – Rev. Brasil. Biol. 59: 693 – 704. Wanntorp, H. E. 1974. Calotropis gigantea (Asclepiadaceae) and Xylocopa tenuiscapa (Hymenoptera: Apidae). – Svensk Bot. Tidsk. 68: 25 – 32. Willis, J. C. and Burkill, I. H. 1903a. Flowers and insects in Great Britain. Part II. – Ann. Bot. 17: 313 – 349. Willis, J. C. and Burkill, I. H. 1903b. Flowers and insects in Great Britain. Part III. – Ann. Bot. 17: 539 – 570. Willis, J. C. and Burkill, I. H. 1908. Flowers and insects in Great Britain. Part IV. – Ann. Bot. 22: 603 – 649. Willson, M. F., Bertin, R. I. and Price, P. W. 1979. Nectar production and flower visitors of Asclepias 6erticillata. – Am. Midl. Nat. 102: 23–35.
Sozialdienst 1,3 Millionen veruntreut – alles fürs Bordell ? Entspannen ist heute Mitarbeiter einer Lebensversicherung vor Gericht / Anwalt: Betrug wurde ihm leicht gemacht bei Büroyoga eingeschränkt sicherungsfachmann, der die peinliche tunterschrift vor, die für die Kassenan- mit der VHS Sache mit Ironie zu überspielen versucht. weisung notwendig war. Allerding
If its consumer rates are frozen, ComEd may actually file for bankruptcy protection, with . Page 1 of 2Story URL: http://news.medill.northwestern.edu/chicago/news.aspx?id=4633 Story Retrieval Date: 8/9/2007 7:40:50 PM CST If its consumer rates are frozen, ComEd may actually file for bankruptcy protection, with broad consequences by Melanie G. Rogers Mar 01, 2007 Commonwealth Ediso
 exposed hosts tended to be more host specific in the
same’’. This conclusion is confirmed by the data that we
tropics, whilst no such pattern was apparent for para-
have presented in this paper. Tropical communities
sitoids utilising hosts concealed in plant tissue. Clearly,
provide some of the best examples of close co-evolved
different categories of species interaction and different
plant-pollinator relationships and in absolute terms do
groups of taxa may or may not show increased special-
contain a higher number of plants with specialised
pollination systems. However, tropical plant assem-
A literature review by Kevan and Baker (1983) con-
blages are on average many times more species-rich than
cluded that ‘‘…from the arctic and alpine areas to the
their temperate counterparts and so may not in fact
lowland tropics, it appears that the frequency of occur-
possess disproportionately more ecologically specialised
rence of specialised pollination syndromes is about the
pollination systems than temperate assemblages.
exposed hosts tended to be more host specific in the
same’’. This conclusion is confirmed by the data that we
tropics, whilst no such pattern was apparent for para-
have presented in this paper. Tropical communities
sitoids utilising hosts concealed in plant tissue. Clearly,
provide some of the best examples of close co-evolved
different categories of species interaction and different
plant-pollinator relationships and in absolute terms do
groups of taxa may or may not show increased special-
contain a higher number of plants with specialised
pollination systems. However, tropical plant assem-
A literature review by Kevan and Baker (1983) con-
blages are on average many times more species-rich than
cluded that ‘‘…from the arctic and alpine areas to the
their temperate counterparts and so may not in fact
lowland tropics, it appears that the frequency of occur-
possess disproportionately more ecologically specialised
rence of specialised pollination syndromes is about the
pollination systems than temperate assemblages.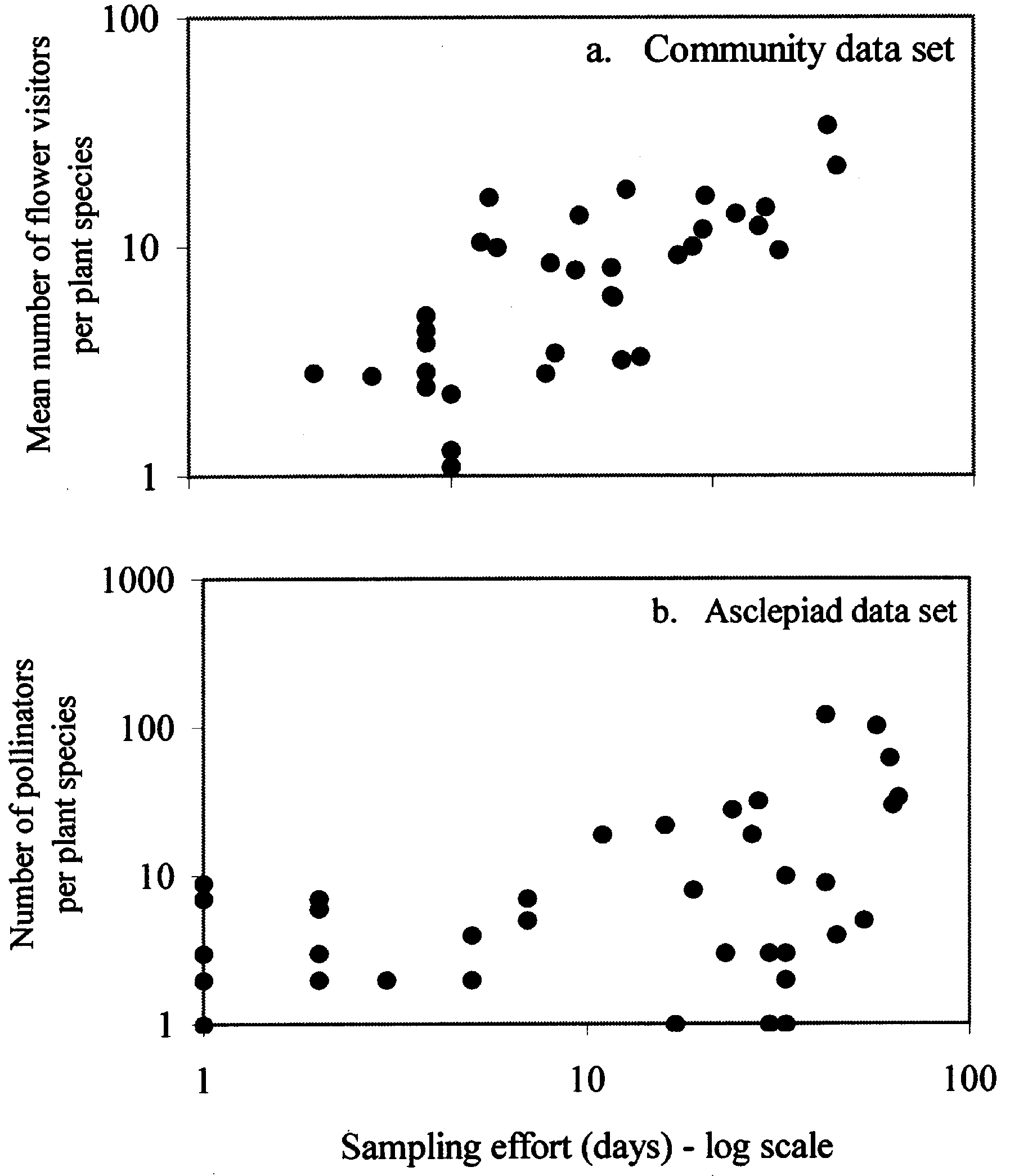 exploring latitudinal trends in plant-pollinator interac-tions, these data sets are as good as any that couldbe currently assembled. We hope that by publishingthis study we will stimulate interest in the question oftropical versus temperate specialisation in ecologicalinteractions and that future researchers will obtaingrants large enough to allow dedicated data collectionthat will tackle this question. Until such time, thesedata sets must suffice.
exploring latitudinal trends in plant-pollinator interac-tions, these data sets are as good as any that couldbe currently assembled. We hope that by publishingthis study we will stimulate interest in the question oftropical versus temperate specialisation in ecologicalinteractions and that future researchers will obtaingrants large enough to allow dedicated data collectionthat will tackle this question. Until such time, thesedata sets must suffice.