Le tadalafil possède une affinité marquée pour la PDE5, mais épargne en grande partie les isoformes PDE1, PDE2 et PDE11, réduisant ainsi le risque d’effets extra-caverneux. L’action se traduit par une augmentation contrôlée de la circulation sanguine locale, indépendante des variations alimentaires. Sa pharmacocinétique repose sur une absorption digestive rapide, un métabolisme hépatique par CYP3A4 et une distribution tissulaire large. La biodisponibilité reste stable, et l’équilibre plasmatique est atteint en quelques jours lors d’administrations répétées. Les interactions cliniquement significatives surviennent avec les inhibiteurs puissants de CYP3A4 tels que le kétoconazole. Dans la littérature pharmacologique, acheter cialis 20 mg est souvent associé à des schémas d’utilisation basés sur la durée prolongée de son action.
Microsoft word - daviesrev0.doc
Antibiotics, present and future
1 Institut Pasteur, Paris, France. 2 University of British Columbia and TerraGen Diversity, Inc., Vancouver BC, Canada
ABSTRACT
The problem of antibiotic resistance in bacterial pathogens is a problem in genetic ecology. For the past half century, microbes have been exposed to enormous quantities of toxicagents (antibiotics) and they have survived. The principal survival strategy employed wasthe functional acquisition and horizontal transfer of antibiotic resistance genes. It isapparent that the genetic mechanisms involved in these processes were novel and that theywere likely to have been involved in the broader aspect of bacterial genome evolution for along period of time. Introduction
Prior to the late 1930s no antibiotics, as we know them, were available. At that timemetal-based disinfectants containing mercury and detergents made from phenoliccompounds were employed and their use may have contributed to present-day resistanceproblems. The antibiotics introduced in the 1940s and thereafter were not new biologicalagents, but rather natural products whose useful antibiotic activity had been discovered;they were naturally present in the environment in small amounts, and genes encodingresistance to them would have been present in the producing organisms and possibly in themicrobial community. The roles of these low-molecular-weight biologically activecomponents in microbial ecology were poorly understood then, as now [8]. What was newand different was the nature and magnitude of their use as a result of theircommercialisation for the treatment of infectious diseases in humans, animals, and plants. Enormous quantities of natural products with demonstrated therapeutic activities(antibiotics) were manufactured for human use by a new generation of pharmaceuticalcompanies and released into the environment, creating a catastrophic situation forterrestrial bacteria. (It has been estimated that more than a million metric tons ofantibiotics have been released into the biosphere during the last 50 years.) What happenedbiologically is a matter of speculation, since microbial ecology was a fledgling science atthe time and retrospective analyses are often difficult to interpret; components of certainlocalisedmicrobial populations may have been wiped out, with significant effects on themicrobial ecology in many different eco-systems, such as changes in species distributionand extensive genetic modification. The use of antibiotics induced an intensive flux ofgenetic information in the microbial world. Essentially a large, uncontrolledmicrobiological ‘experiment’ took place world-wide in the second half of the 20th century,with only the survivors available for investigation. Retrospective analyses of this‘experiment’, properly executed and interpreted, give important information on subjectssuch as horizontal gene transfer and genome evolution. Microbial Biosystems: New Frontiers Proceedings of the 8th International Symposium on Microbial Ecology Bell CR, Brylinsky M, Johnson-Green P (eds) Atlantic Canada Society for Microbial Ecology, Halifax, Canada, 1999. Development of antibiotic resistance
With few exceptions antibiotic resistance in bacterial pathogens was identified veryquickly after the introduction of antibiotics into clinical practice. While mutations leadingto resistance occurred extensively (for example, streptomycin resistance inM. tuberculosis), it is likely that mutations leading to low levels of resistance were only a‘first’ step in the development of the refractory strains found today [9]. However, themajority of bacteria acquired the genes encoding resistance to antibiotics from exogenousand still largely unidentified sources. Obviously only infection-related strains (the tip ofthe iceberg) were identified and studied; it is unfortunate that changes in antibioticsusceptibility patterns of microbial populations could not have been analysed at earliertimes and even more disappointing that the demographics of microbial populations underantibiotic stress have been little studied since! It would have been of great value to tracethe course of genetic traffic on the introduction of antibiotics. Did resistance determinantsdevelop in many species or in only a limited number, before being mobilised andtransferred to the pathogens identified as antibiotic-resistant? This may never be known. However, it is to be hoped that the next time a completely novel antibiotic (chemicalentity) is introduced into human clinical practice, steps will be taken to ensure analysis andelucidation of the route(s) by which antibiotic resistance can be established in microbialcommunities.
Prior to the introduction and use of antibiotics, antibiotic-resistant microbes were absent
from human or animal flora [7, 11]. With antibiotic use, rapid increases in bacterialresistance to tetracycline and other antibiotics was documented in previously susceptiblestrains. Recent examples come from studies of antibiotic-resistance development in E. coli0157:H7 in the State of Washington: in 1984 all strains were antibiotic-sensitive, in 19897% of the isolates carried antibiotic-resistance determinants, and since that time thenumber and incidence of resistance characters has expanded. The dramatic appearance ofantibiotic resistance in pathogenic Shigella spp. during an epidemic of intestinal infectionsin the 1950s in Japan is yet another example of the rapid response of bacterial pathogens tothe threat of antibiotic use [17] and signalled the appearance of resistance plasmids.
At the present time many different resistance genes (often termed ‘cassettes’) exist in
accessible and mobilisable form in the microbial gene pool. Upwards of a dozenbiochemical mechanisms of resistance have been characterized (Table 1): these areencoded by hundreds of different genes, many of them identified as classes of relatedallelic determinants. The bacterial population has essentially stockpiled a considerablearmamentarium of genetic defences. Not surprisingly the majority of resistance genes arecarried by plasmids, transposons, and other elements capable of inter-generic andinter-specific mobility. While information on the origins and acquisition of these genes ishard to come by (where did they come from and how did they move to where they are?),significant advances have been made. Origins of antibiotic-resistance genes
As with the pathogenicity islands found in clinical isolates, resistance genes are oftenclustered on their resident genomes and differ in base composition from their immediatesequence environment. No single source of resistance genes can be identified but anumber have been proposed, including housekeeping genes (alternate substrates, mutationto new substrate recognition, etc.), antibiotic-producing microbes (self-protection), and
Table 1: Biochemical mechanisms of antibiotic resistance and their genetic determinants Genetic determinants Mechanism Examples Mutation Gene acquisition Table 2: Resistance determinants with biochemical homologues in antibiotic-producing organisms Antibiotic Resistance mechanisms*
acetyltransferases, phosphotransferases, adenyltransferases
*Multiple drug resistance (MDR) efflux systems are common to producing organisms and clinical isolates
‘natural’ resistance genes in soil communities. Supporting evidence is largelycircumstantial and comes from a variety of studies. In no case can a direct (single transfer)relationship be established between the resistance gene(s) in clinical isolates and theputative source organisms. For example, methicillin resistance (the mec cluster) found inStaphylococcus aureus is closely related to that of Staphylococcus sciuri [24], but transferin vivo has not been shown. The enterobacterial aminoglycoside acetyltransferase (aac)genes, some of which are derived from chromosomal genes in gram-negative bacteria [13],are another example; interestingly the aac genes have been shown to be members of alarge protein N-acyltransferase family [18].
That resistance to aminoglycosides, macrolides, tetracycline, chloramphenicol, and
other antibiotics may have originated in their producing organisms is indicated bybiochemical and nucleic acid sequence similarities of the genes and their products (Table2). An example of an antibiotic-resistance gene of unusual parenthood is provided bymupirocin (pseudomonic acid), an antibiotic employed in the treatment of topicalinfections by gram-positive bacteria. Resistance to mupirocin is due to the presence of analtered isoleucyl tRNA synthase that may have originated in eukaryotes and was onlyrecently acquired by S. aureus strains [5]. The molecular details of this unusualacquisition remain to be ascertained. Mutation
Extensive ‘tailoring’ and adaptation of the different genes must occur during transferbetween the ‘source’ organisms and the ultimate clinical isolates, which are usuallycharacterised by a different G + C composition, codon usage pattern, and regulatoryelements. How do these modification occur? It is probable that uncharacterisedintermediate transfer hosts play a role in the tailoring of the determinants currentlyidentified. Hyper-mutable strains and mutator genes must have played roles, and recentstudies have shown that many natural bacterial isolates, particularly pathogenic strains, arehyper-mutagenic and carry mutator genes (mutS, mutL, mutT) [14, 21]. In some cases themutator genes involved, such as mutT, favour the reduction of G/C-rich sequences,contributing to adaptation of codon usage patterns to new cellular environments [20]. Thenatural incidence of mutator strains in the environment and their potential roles in theevolution of antibiotic resistance genes (or any horizontally transferred determinants, suchas biodegradation clusters) can only be hypothesized. The critical juxtaposition ofmutational and gene transfer events is not understood, apart from examples such as theevolution of extended ß-lactamases by natural protein engineering [6]. In addition, thegenetic ecology of antibiotic resistance is inextricably linked to the ecology of microbialpopulations in humans, animals, and plants. Mechanisms of acquisition (resistance-gene capture)
The tandem assembling of multiple drug-resistance genes in a single, mobile geneticelement has many implications:(a) A single plasmid replicon or transposon may carry resistance to a number ofclinically significant antibiotics at the same time. (b) A single gene may encode a biochemical mechanism that engenders resistance to anentire class of antimicrobials, e.g. all available macrolides (erm), all ß-lactams (bla), orseveral different aminoglycosides (aac, aph, ant).
(c) A single resistance gene may encode resistance to at least two structurally unrelatedantibiotics, e.g. streptomycin and spectinomycin (aad). (d) Use of a non-antibiotic substance, such as a disinfectant or metal salt, may select forlinked antibiotic-resistance genes carried on a plasmid or transposon. The most common mechanism by which tandem arrays of antibiotic-resistance genes areassembled in the Enterobacteriaceae is gene-capture by integrons. This process has beendissected genetically and biochemically by Roy, Sundstrom, Hall and Stokes, and their co-workers[4, 10, 12]. All known integrons are composed of three essential elements: anintegrase gene, a primary recombination site and a strong promoter. The integron-associated integrases belong to the site-specific recombinase family and are able torecombine with discrete units of DNA, the gene cassettes, providing them with a promoterfor their expression. The resistance cassettes contain a single resistance gene associatedwith a specific recombination sequence, known as the 59-base elements. An outline of theintegron-integrase acquisition process is illustrated in Fig. 1. Many important details ofthis process need to be elucidated. For example, what is the origin of the integrases andthe 59-base-pair elements which are essential for the recognition and integration of thecaptured gene cassettes? Are they derived from bacteriophages? What is the origin of thegene cassettes and how do the 59-base-pair elements become attached?
Recent work by Mazel et al. on the antibiotic-resistance integrons and the Vibriocholerae repeated sequences (VCRs) [16] has demonstrated striking similarities, whichsuggests important functional relationships between these compound structures. Both theVCR and antibiotic-resistance integrons possess specific integrases that insert codingsequences (ORFs)into a unique chromosomal attachment site, leading to the formation oftandem arrays of genes. In the case of V. cholerae the cluster of VCR-associated ORFsrepresents 60-100 genes and occupies 4% of the 2.5Mb genome [3]. Preliminary studiesindicate that such integrase-linked structures are found in a number of different Vibrio spp. The VCR integrases so far identified form a class related to, but distinct from, IntI2 (ofclass II integrons).
The VCR structures differ from the integrons in that (1) a defined integration site is
lacking, (2) a certain number of the ORFs possess their own promoter, (3) an identicalrepeat (VCR) is associated with each ORF, (4) the ORFs encode largely unknownfunctions; those so far identified appear to be related to virulence, and no well-definedantibiotic-resistance genes are in the clusters, although it should be noted that the blaP anddfrVI cassettes of integrons are VCR-associated. Thus resistance and virulence are linked,which implies that the selective pressure of antibiotic use played a part in thedissemination of pathogenicity determinants in the development of ‘new’ pathogens. Many Vibrio spp. possess similar clusters, and detailed analysis of the open reading frames(in process) confirms the diversity of sequences present. If each Vibrio species possessesVCR-like clusters of hundreds of unidentified genes, a veritable treasure trove will beavailable for functional genomic studies. The number of different bacterial integrons maybe very large, each classified according to its associated site-specific integrase. Integron-driven gene capture is likely to play an important role in the evolution of bacterial genomesbeyond the functions of antibiotic resistance and pathogenicity. While such tandem arrayshave been identified only in enteric bacteria, incomplete integrons have been found in thegram-positive M. tuberculosis and Corynebacterium glutamicum, and clusters of resistancegenes have been characterised in Staphylococcus aureus [1]. Fig. 1: The mechanism by which resistance-gene cassettes are incorporated into capture elements such as integrons or VCRs (see text). The gene intI1 encodes the integrase (Int) and the promoter (Pant) necessary for transcription of inserted downstream genes. The direction of transcription of the int gene is shown by the lower arrow. The resistance gene cassettes (ant1R, 2R) are integrated at the attachment site (attI) which interacts with the 59-base-pair elements (solid bar) in each resistance-gene cassette. The big picture – the ecology of antibiotic-resistant bacteria
We have focused on the acquisition, dissemination and organisation of genes within thebacterial kingdom: the genetic ecology of antibiotic resistance. On the other hand, thewide distribution of antibiotic-resistant bacteria in many different ecological niches definesthe macro-ecology of infectious disease; the spread of antibiotic-resistant organisms aswell as the horizontal transfer of resistance genes must be considered. Antibioticresistance is associated with many different sources, and the following are importantfactors:a)
health care institutions and the maintenance and transmission of resistant strains
within the community (food, domestic animals, etc.)b)
the dissemination of antibiotic-resistant bacteria in animals and their different
global transfer of antibiotic-resistant strains (pathogens and non-pathogens) due to
increased international travel and the food ‘business’, including traffic in live animals andassociated products.
The components of the food chain are inextricably linked and interactions between
bacterial populations of humans, animals, plants and environment disseminate antibiotic-resistant pathogens within the food chain. The threat of serious infection from food-bornepathogens has become a significant risk to the consumer [15]. The implications for humanhealth, especially among disease-susceptible groups such as the very young and theelderly, must be addressed as a priority. Typically, antibiotic usage in agriculture at firstled to transfer of resistance genes from animal isolates (such as Salmonella typhimurium29] to human pathogens. However, the increasing incidence of multidrug-resistant strainsof S. typhimurium DT104 and E. coli 0157 H7 [2, 22, 23], which are pathogenic to bothanimals and humans, is a different matter and is cause for concern [19]. Conclusions
A general understanding of the origins, mechanisms of acquisition and dissemination ofantibiotic-resistance genes within microbial populations exists currently. However, thereare many missing links in this picture. Studies of antibiotic-resistant bacteria have focusedon the properties of the antibiotic-resistant pathogens isolated in human clinical situations,with little effort expended in trying to establish their genesis. Such information will becritical in providing the means to control the problem of antibiotic resistance in microbes. Efforts to reduce the development and incidence of antibiotic resistance in hospitals andthe community must be increased on a world-wide basis. The introduction of novelantibiotics and effective vaccines is a priority, as is improved technology in the agriculturaland food supply industries that will reduce dependence on antibiotic use and the exclusionof resistant organisms from the food chain. Nationwide and international surveillance willbe needed to identify outbreaks and to institute effective measures in containment. Acknowledgements
This work was supported by the Canadian Bacterial Diseases Network, the NationalScience and Engineering Council (Canada) and the Institut Pasteur, Paris. We are gratefulto Dorothy Davies for assistance in preparing the manuscript. References
1. Allignet J, Aubert S, Morvan A, El Solh N (1996) Distribution of genes encoding
resistance to streptogramin A and related compounds among staphylococci resistant tothese antibiotics. Antimicrob Agents Chemother 40:2523-2528.
2. Armstrong GL, Hollingsworth J, Morris JG, Jr (1996) Emerging foodborne pathogens:
Escherichia coli 0157:H7 as a model of entry of a new pathogen into the food supplyof the developed world. Epidemiol Rev 18:29-51.
3. Barker A, Clark CA, Manning PA (1994) Identification of VCR, a repeated sequence
associated with a locus encoding a hemagglutinin in Vibrio cholerae O1. J Bacteriol176:5450-5458.
4. Bissonnette L, Roy PH (1992) Characterization of In0 of Pseudomonasaeruginosa
plasmid pVS1, an ancestor of integrons of multiresistance plasmids and transposons ofgram-negative bacteria. J Bacteriol 174:1248-1257.
5. Brown JR, Zhang J, Hodgson JE (1998) A bacterial antibiotic resistance gene with
eukaryotic origins. Curr Biol 8:R365-367.
6. Bush K, Jacoby GA, Medeiros AA (1995) A functional classification scheme for b-
lactamases and its correlation with molecular structure. Antimicrob Agents Chemother39:1211-1233.
7. Datta N, Hughes VM (1983) Plasmids of the same Inc groups in Enterobacteria before
and after the medical use of antibiotics. Nature 306:616-617.
8. Davies J (1990) What are antibiotics? Archaic functions for modern activities. Mol
9. Davies J, Webb V (1998) Antibiotic resistance in bacteria. In: Krause RM (ed)
Emerging Infections. Academic Press, New York, pp. 239-273
10. Hall RM, Collis CM (1995) Mobile gene cassettes and integrons: capture and spread of
genes by site-specific recombination. Mol Microbiol 15:593-600.
11. Hughes VM, Datta N (1983) Conjugative plasmids in bacteria of the 'pre-antibiotic'
12. Huovinen P, Sundström L, Swedberg G, Sköld O (1995) Trimethoprim and
sulfonamide resistance. Antimicrob Agents Chemother 39:279-289.
13. Lambert T, Gerbaud G, Courvalin P (1994) Characterization of the chromosomal
aac(6') -Ij gene of Acinetobacter sp. 13 and the aac(6')-Ih plasmid gene ofAcinetobacter baumannii. Antimicrob Agents Chemother 38:1883-1889.
14. LeClerc JE, Li B, Payne WL, Cebula TA (1996) High mutation frequencies among
Escherichia coli and Salmonella pathogens. Science 274:1208-1211.
15. MAFF Report. 1998. A review of antimicrobial resistance in the food chain. Ministry
of Agriculture, Fisheries and Food, London UK.
16. Mazel D, Dychinco B, Webb VA, Davies J (1998) A distinctive class of integron in the
Vibrio cholerae genome. Science 280:605-607.
17. Mitsuhashi S (1993) Drug resistance in bacteria: history, genetics and biochemistry. J
18. Neuwald AF, Landsman D (1997) GCN5-related N-acetyl-transferases belong to a
diverse superfamily that includes the yeast SPT10 protein. Trends Biochem Sci22:154-155.
19. Perreten V, Schwarz F, Cresta L, Boeglin M, Dasen G, Teuber M (1997) Antibiotic
resistance spread in food. Nature 389:801-802.
20. Taddei F, Hayakawa H, Bouton M-F, Cirinesi A-M, Matic I, Sekiguchi M, Radman M
(1997) Counteraction by MutT protein of transcriptional errors caused by oxidativedamage. Science 278:128-130.
21. Taddei F, Matic I, Godelle B, Radman M (1997a) To be a mutator; or how pathogenic
and commercial bacteria can evolve rapidly. Trends Microbiol 5:427-428.
22. Threfall EJ, Frost JA, Rowe B (1996) Increasing spectrum of resistance in
multiresistant Salmonella typhimurium. Lancet 347:1053-1054.
23. Wall PG, Morgan D, Lamden K, Griffin M, Threlfall EJ, Ward LR, Rowe B (1995)
Transmission of multi-resistant strains of Salmonella typhimurium from cattle to man. Vet Rec 136:591-592.
24. Wu S, Piscitelli C, de Lencastre H, Tomasz A (1996) Tracking the evolutionary origin
of the methicillin resistance gene: cloning and sequencing of a homologue of mecAfrom a methicillin susceptible strain of Staphylococcus sciuri. Microb Drug Resistance2:435-441.
EUNHEE EMILY KO 1630 Chicago Ave Apt.707 Evanston, IL 60201 EunKo2013@u.northwestern.edu 814-321-5088 Web Site: eunheeko.com ~ QUANTITATIVE MARKETING PROFESSIONAL ~ Dedicated professional with a commitment to driving profitability through skillful quantitative modeling and in-depth analysis of market, competitor, industry and economic insights. Excellent background include
International Journal of Phytomedicine 2 (2010) 363-372 Research article Phytoestrogens of Pachyrhizus erosus prevent Bone Loss in an Ovariectomized Rat Model of Osteoporosis Arief Nurrochmad1*, Fransiska Leviana2, Caecilia Govita Wulancarsari3, Endang Lukitaningsih4 * Corresponding author: Abstract The effects of the etyl acetate extract of root of Pachyrhizus A
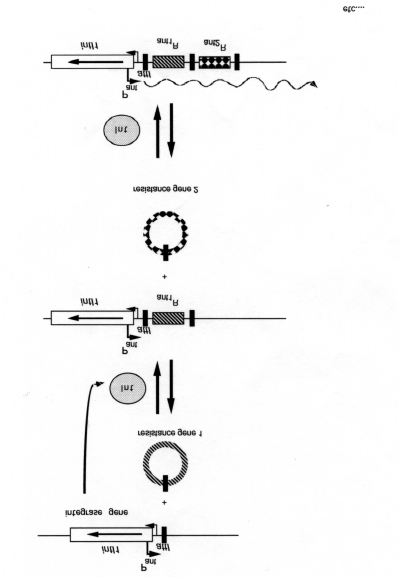 Fig. 1: The mechanism by which resistance-gene cassettes are incorporated into capture elements such as
Fig. 1: The mechanism by which resistance-gene cassettes are incorporated into capture elements such as