Le tadalafil possède une affinité marquée pour la PDE5, mais épargne en grande partie les isoformes PDE1, PDE2 et PDE11, réduisant ainsi le risque d’effets extra-caverneux. L’action se traduit par une augmentation contrôlée de la circulation sanguine locale, indépendante des variations alimentaires. Sa pharmacocinétique repose sur une absorption digestive rapide, un métabolisme hépatique par CYP3A4 et une distribution tissulaire large. La biodisponibilité reste stable, et l’équilibre plasmatique est atteint en quelques jours lors d’administrations répétées. Les interactions cliniquement significatives surviennent avec les inhibiteurs puissants de CYP3A4 tels que le kétoconazole. Dans la littérature pharmacologique, acheter cialis 20 mg est souvent associé à des schémas d’utilisation basés sur la durée prolongée de son action.
Storedproducts.okstate.edu
Attractancy and Toxicity of an Attracticide for Indianmeal Moth, Plodia interpunctella (Lepidoptera: Pyralidae)
CHRISTIAN NANSEN1 AND THOMAS W. PHILLIPS2
ABSTRACT Plodia interpunctella (Hu¨bner) is a serious and widespread postharvest pest on cereal
products, dried fruits, candy, and pet food. Due to the strong positive anemotactic ßight response of
P. interpunctella males to the main component of the female-produced pheromone [(Z,E)-9,12-
tetradecadienyl acetate, herein referred to as ZETA], we evaluated the potential of an attracticide for
this pest, in which ZETA as attractant was combined with permethrin as the killing agent. Two
concentrations of ZETA [0.16 and 0.32% (wt:wt)] and Þve concentrations of permethrin [0, 3, 6, 12,
and 18% (wt:wt)] were incorporated into Last Call gel (10 different permethrin:ZETA combinations).
All attracticide gels were evaluated in a toxicity test, in which either the tip of a leg or an antenna of
a virgin P. interpunctella male was touched Ͻ3 s into a dot of attracticide gel. These males were
subsequently transferred to jars with virgin females. The toxicity test showed that a brief and gentle
contact of P. interpunctella males with attracticide gel containing 3Ð18% permethrin caused a signif-
icant reduction in mating and killed males moths within 24 h. A wind tunnel test was conducted to
evaluate the ßight responses of P. interpunctella males to the same 10 attracticide gels. Male moths
displayed signiÞcantly higher levels of positive anemotactic ßight and more males made contact with
the attracticide gel when the ZETA concentration was 0.16% compared with 0.32%. P. interpunctella
males showed no signs of repellency to permethrin concentrations within a range of 0Ð18% in the
attracticide gel. Three densities of P. interpunctella pairs were released into small warehouse rooms,
and we found that the attracticide gel suppressed oviposition when the moth density was at a low level,
but it was ineffective when the moth density exceeded one male-female pair per 11.3 m3. KEY WORDS pheromone, stored-products, permethrin, attract-and-kill, oviposition
THE INDIANMEAL MOTH, Plodia interpunctella (Hu¨bner)
Pheromone-baited sticky traps are widely used for
(Lepidoptera: Pyralidae), is a widespread and serious
early detection and monitoring of stored-product
pest on bulk-stored grain and seeds (Lecato 1976,
moth populations in and around food facilities, and the
Storey et al. 1983, Vick et al. 1986, Cuperus et al. 1990,
main component of the P. interpunctella female-pro-
Doud and Phillips 2000, Nansen et al. 2004), ßour, feed
duced pheromone [(Z,E)-9,12-tetradecadienyl ace-
and other milled products (Lecato 1976), prototype
tate (Z9,E12Ð14:Oac, herein referred to as ZETA)]
military rations (Cline and Highland 1985), dried
was identiÞed by Brady et al. (1971) and Kuwahara et
fruits (Johnson et al. 1992), and nuts (Johnson et al.
al. (1971), and ZETA was among the Þrst pheromones
1992). Damage of food products is attributed to feed-
to become commercially available (Phillips 1997).
ing by larvae, which are capable of penetrating a wide
Later, three additional components of the P. inter-
range of packing materials (Cline 1978). Infestations
punctella female-produced pheromone were identi-
by P. interpunctella can have a great economic impact
Þed (Kuwahara and Casida 1973, Sower et al. 1974,
due to direct product loss and indirectly to factors
Soderstrom et al. 1980, Teal et al. 1995, Zhu et al. 1999)
such as the cost of pest control and loss of sales from
in the following relative ratio to 100 U of ZETA (Zhu
consumer complaints (Phillips et al. 2000a). The fu-
et al. 1999): 1) (Z,E)-9,12-tetradecadienal (Z9,E12Ð
migant methyl bromide is widely used to control P.
14:Ald), 11 U; 2) (Z,E)-9,12-tetradecadienol (Z9,E12Ð
interpunctella and other stored-product pests in food
14:OH), 18 U; and 3) (Z)-9-tetradecenyl acetate (Z9Ð
processing and warehousing facilities, and the antic-
14:Oac), 12 U. However, most commercial P.
ipated U.S. ban in 2005 of this fumigant has motivated
interpunctella lures only contain ZETA. The response
research on alternative methods to control stored-
of male moths to female-produced sex pheromone
product insect pests (Fields and White 2002).
may also be used for suppression of these insect pests
through either mating disruption or an attracticide
1 Department of Entomology, Leon Johnson Hall, Montana State
(Phillips et al. 2000b). Mating disruption involves re-
University, Bozeman, P.O. Box 173020, MO 59717 (e-mail:
lease of sex pheromone in high amounts to confuse
2 Department of Entomology and Plant Pathology, 127 Noble Re-
male moths in their search for conspeciÞc females and
search Center, Oklahoma State University, Stillwater, OK 74078.
thereby reduce the frequency of mating (Carde´ and
0022-0493/04/0703Ð0710$04.00/0 ᭧ 2004 Entomological Society of America
Minks 1995). Mating disruption for stored-product
ducted with virgin adults that were 1Ð2 d old; adults
moths has been evaluated for Sitotroga cerealella
(Olivier) (Vick et al. 1978), Ephestia cautellaAttracticide. A commercially developed attracti-
(Walker) (Mafra-Neto and Baker 1996, Shani and
cide gel formulation known as Last Call was used and
Clearwater 2001), and P. interpunctella (Ryne et al.
prepared for us as needed by the manufacturer (IPM
2001). An attracticide involves combination of an at-
Technologies Inc., Portland, OR). We examined Þve
tractant, typically a sex pheromone and/or a food
concentrations of permethrin [0, 3, 6, 12, and 18%
odor, and a killing agent (pathogen or pesticide), so
(wt:wt)] and two concentrations of ZETA [0.16 and
that the insects are attracted to a point source and
0.32% (wt:wt){rsqb] (total of 10 attracticide gels). In
subsequently killed after contact with the attracticide
all experiments, we used individual attracticide gel
(Lanier 1990). Compared with traditional applications
of pesticides, the main advantages of using an attrac-
Toxicity Test. Individual P. interpunctella males
ticide are 1) insecticides are not broadcast over large
were held with “feather weight” forceps (BioQuip,
areas; 2) insecticides are not applied directly onto
Rancho Dominguez, CA), and a single leg or antenna
food materials; 3) nontarget insects (beneÞcials) are
was dipped gently into a dot of attracticide for Ͻ3 s.
unlikely to be affected; and 4) although relatively high
All 10 attracticide gels were examined, and 10 P. in-
concentrations may be used in point sources, the total
terpunctella males were tested individually for each
amount of pesticide needed to treat a facility is con-
attracticide gel. Immediately after the forced contact
siderably smaller overall compared with a broadcast
with the attracticide, each male was transferred to a
treatment. EfÞcient insect control based on attracti-
250-ml glass jar containing 15 g of whole wheat kernels
cides has been shown for a number of important moth
and a virgin P. interpunctella female. After 24 h, the
pests on forest trees [western spruce budworm, Cho-
whole wheat kernels were sifted, the number of eggs
ristoneura occidentalis Freeman (Sower and Shorb
laid was counted, and the status of the male was eval-
1985)], Þeld crops [pink bollworm, Pectinophora gos-
uated and assigned a score as follows: dead, 0; immo-
sypiella (Saunders) (Haynes et al. 1986, Miller et al.
bile but alive, 1; and alive, 2. Virgin females lay eggs if
1990); Egyptian cotton leafworm, Spodoptera littoralis
they are kept for 4Ð5 d (C.N., unpublished data), so a
Boisduval (De Souza et al. 1992, Downham et al.
control group of 10 jars with wheat was set up in which
1995)], and fruits [navel orangeworm, Amyelois tran-
a single virgin female was kept under the same con-
sitella (Walker) (Phelan and Baker 1987), codling
ditions for 24 h without introducing a P. interpunctella
moth, Cydia pomonella (L.) (Charmillot and Hofer
1997), and light brown apple moth, Epiphyas postvit-Wind Tunnel Experiment. The ßight response of P. tana (Walker) (Brockerhoff and Suckling, 1999, Suck-
interpunctella males to the attracticide was examined
ling and Brockerhoff 1999)]. Trematerra and Capizzi
in a wind tunnel (square cross section of 0.9 by 0.9 m,
(1991) described the use of an attracticide to control
1.8 m in length), typical of those used for moth ßight
the Mediterranean ßour moth, Ephestia kuehniella
bioassays (Haynes and Baker, 1989). An aluminum
mesh screen was mounted in front of the fan to keep
In this study, we evaluated a commercially formu-
insects inside the wind tunnel. Room air entered the
lated attracticide for P. interpunctella in which ZETA
wind tunnel through a screen Þlter impregnated with
was used as the attractant and the synthetic pyrethroid
activated carbon at the upwind end, and air was pulled
permethrin was the killing agent. Experiments were
through the tunnel and exhausted out of the room by
designed to determine 1) to what extent subtle contact
an electric fan mounted equidistant from the tunnel
of P. interpunctella males with the attracticide would
sides in a sheet-metal reduction plenum. The wind
affect their survival and ability to mate; 2) the positive
tunnel and air in the room housing it were kept at
anemotactic ßight response of P. interpunctella males
18Ð22ЊC and 30Ð40% RH, respectively. The airßow
to the attracticide in a wind tunnel; and 3) the po-
rate inside the tunnel was Ϸ10 cm/s. A 10 by 10-cm
tential of the attracticide to suppress P. interpunctella
platform was placed 30 cm from the ßight tunnel ßoor
populations under controlled, simulated warehouse
in the upwind end and was used to hold a microscope
conditions with different moth densities.
slide with a dot of attracticide gel, which was intro-
duced into the wind tunnel 15 min before the ßight
response of the Þrst P. interpunctella male was evalu-
Materials and Methods
ated. Attracticide gel dots were replaced after every
four to Þve trials. In the downwind end of the wind
Insects. P. interpunctella adults from the laboratory
tunnel 30 cm from the ßoor in front of the fan, indi-
culture at Oklahoma State University were reared in
vidual P. interpunctella males were released from a
a growth chamber at a photoperiod of 16:8 (L:D) h,
cylindrical screened release cage (5 cm in diameter
28ЊC, and 60Ð70% RH on a standardized diet of corn
and 4 cm in height), which was placed on a metal
meal, chick laying mash, chick starter mash, and glyc-
erol at a volumetric ratio of 4:2:2:1, respectively. The
P. interpunctella males were tested individually, and
light program of the growth chamber was set so that
they were kept inside the release cage for precondi-
8 a.m. corresponded to the beginning of the scoto-
tioning for 5 min before release. Experiments were
phase. Pupae were sexed and carefully transferred to
conducted under dim lighting during morning and
individual 2-ml glass vials. All experiments were con-
afternoon hours. The following behavioral responses,
NANSEN AND PHILLIPS: ATTRACTICIDE FOR P. interpunctella
similar to those used by Haynes et al. (1986), were
were changed randomly among the storage sheds, and
recorded: 1) wing fanning in the release cage during
compressed air was used to clean the walls inside the
preconditioning was scaled from 0 to 3 with 0 being no
storage sheds between subsequent trials. Each trial
fanning and three being intensive fanning; 2) take-off
lasted 76 h and was repeated six times. Pairs of moths
(yes/no); 3) time spent at release platform before
were released in three densities (females:males): 1:1,
take-off; 4) positive anemotactic ßight (yes/no); 5)
landing on the platform holding the attracticide (yes/
Statistical Analysis. The PROC MIXED procedure
no); 6) time of ßight from release until landing on the
with inbuilt contrasts in PC-SAS 8.0 (SAS Institute
platform; 7) direct contact with the attracticide dot
1999) was used to examine the concentration effects
(yes/no); and 8) the type of contact with the attrac-
of ZETA and permethrin on 1) the ovipositional re-
ticide was scaled from 0 to 3 of P. interpunctella males
sponse of P. interpunctella females mated to males in
that landed on the platform: 0, no touch; 1, gentle
the toxicity test; 2) the behavioral ßight response by
touch with either a single leg or with just one antenna;
males in the wind tunnel experiment; and 3) the ovi-
2, touch with at least two body parts (e.g., two legs, one
positional response (total number of eggs laid in the
leg, and an antenna) or touch with a wing; and 3, when
three food dishes per number of P. interpunctella fe-
the male either crossed the attracticide gel dot by
males) in the suppression study in the simulated ware-
walking over it, got stuck in the gel, or in other ways
houses. The PROC NPAR1WAY in SAS was used to
had distinct or prolonged contact with the attracticide
conduct a KruskalÐWallis test with Wilcoxon scores of
gel. A trial was terminated within a maximum of 15
the concentration effects of ZETA and permethrin on
min. We evaluated the ßight response of 7Ð10 P. in-
1) ranked male status after 24 h in the toxicity test; 2)
terpunctella males for each attracticide gel on two
ranked observations of fanning in the release cage, 3)
separate days (a total of 15Ð20 males per attracticide
binomial data on upwind ßight response and landing
gel). A weak soap solution was used to clean the wind
on platform containing the attracticide; and 4) ranked
tunnel between bioassays with different attracticide
observations of the male contact with dots of the
Suppression Study. Three metal storage sheds
(Piedmont, Mauldin, SC), each of 11.3 m3 [2.3 m
(width) by 3.0 m (length) by 1.7 m (height)], were
used as simulated warehouses for evaluation of the
attracticide gel. The efÞcacy of the attracticide gel was
Toxicity Test. P. interpunctella females laid no or
determined by comparing oviposition by P. interpunc-
very few eggs during 24 h when no male was offered
tella in three storage sheds with the following simul-
in control experiments (Fig. 1). In the PROC MIXED
taneous treatments: 1) one freely exposed attracticide
analysis of eggs laid, we found that forced male moth
gel dot was applied to a paper card placed on the
contact with permethrin had a signiÞcant effect on
center of the south wall in the storage shed; 2) an
total oviposition by females (F4,90 ϭ 4.89; P Ͻ 0.01),
attracticide gel dot was similarly applied to the south
whereas ZETA concentration had no signiÞcant effect
wall in another storage shed, but a screen cage
on the total oviposition (F1,90 ϭ 0.27; P ϭ 0.61). There
(1.2-mm mesh opening) was mounted around the
was no signiÞcant interaction effect between per-
attracticide to allow the males only to come within a
methrin concentration and ZETA concentration (F4,90
2-cm range of the gel dot; and 3) no gel dot (control).
ϭ 1.26; P ϭ 0.29). Because there was no signiÞcant
Oviposition was assessed by counting the total number
effect in toxicity caused by ZETA concentration, we
of eggs laid by P. interpunctella females in three petri
grouped the results from 0.16 and 0.32% ZETA and
dishes in each storage shed (10 cm in diameter), each
contrasted oviposition results across permethrin con-
containing 10 g of whole wheat kernels treated with
centrations (Fig. 1). There was no signiÞcant differ-
walnut oil (10 l/g wheat kernel). Nansen and Phillips
ence in oviposition when males had been exposed to
(2003) determined that whole wheat kernels treated
attracticides containing 0 or 3% permethrin (F1,90 ϭ
with this concentration of walnut oil stimulated ovi-
1.07; P ϭ 0.30), but total oviposition was signiÞcantly
position by P. interpunctella females. The three food
lower when attracticides contained 6% permethrin
dishes were placed on a wooden shelf (60 by 120 cm)
compared with 3% (F1,90 ϭ 6.31; P ϭ 0.01). In the
suspended in the center of the storage shed 70 cm
KruskalÐWallis test of the ranked male status after
above the ground. The suppression studies were con-
24 h, we found that permethrin concentration in at-
ducted during winter (December 2002ÐApril 2003),
tracticides had a signiÞcant effect on males status after
and an electric heater was placed underneath the
24 h (2 ϭ 64.04, df ϭ 4, P Ͻ 0.01), whereas ZETA
wooden shelf in each of the three storage sheds to
concentration (2 ϭ 0.22, df ϭ 1, P ϭ 0.64) did not
maintain the temperature between 25 and 30ЊC and
have signiÞcant inßuence on the male status after 24 h
30Ð40% RH (monitored with Hobo data logger in each
(Fig. 2). Again, we grouped the results from 0.16 and
storage shed). Based on the results from the toxicity
0.32% ZETA and made paired comparisons of the
test and the wind tunnel experiment, we evaluated the
malesÕ status at different permethrin concentrations.
attracticide gel containing the combination of 6% per-
There was a signiÞcant difference in male status of
methrin and 0.16% ZETA in the suppression study.
treatments with 0 and 3% permethrin (2 ϭ 6.84, df ϭ
The suppression study was conducted in complete
1, P Ͻ 0.01), and 3 and 6% permethrin (2 ϭ 15.7, df ϭ
darkness. For each replication, the three treatments
1, P Ͻ 0.01), but there was no signiÞcant difference in
Fig. 1. Mean oviposition within 24 h of single P. interpunctella couples after the male had subtle contact with one of 10
attracticides containing 0.16 or 0.32% ZETA and 0, 3, 6, 12, and 18% permethrin. Results with different ZETA concentrations
were grouped because only permethrin concentration had signiÞcant effect on oviposition (as indicated by different letters).
Oviposition by females not paired with a male was not included in the statistical analysis.
male status for treatments with attracticides contain-
more P. interpunctella males performed upwind ßight
ing 6, 12, or 18% permethrin (P Ͼ 0.05).
toward attracticide gels containing 0.16% ZETA (n ϭ
Wind Tunnel Experiment. The ßight responses of
77, 0.83 Ϯ 0.04 [SE]) than to those containing 0.32%
P. interpunctella males in the wind tunnel experiment
(n ϭ 74, 0.63 Ϯ 0.06) (df ϭ 1, 2 ϭ 7.40, P Ͻ 0.01), but
are summarized in Table 1. Most of the tested P.
the proportion of males performing upwind ßight was
interpunctella males performed considerable fanning
not affected signiÞcantly by the permethrin concen-
during the preconditioning, and neither permethrin
tration (2 ϭ 5.52, df ϭ 4, P ϭ 0.24) (Table 1). The
concentration (2 ϭ 8.26, df ϭ 4, P ϭ 0.22) nor ZETA
proportion of upwind-ßying P. interpunctella males
concentration (2 ϭ 0.13, df ϭ 1, P ϭ 0.72) had sig-
landing on the platform with the attracticide was not
niÞcant effect on the scoring of fanning. All P. inter-
affected by the permethrin concentration (2 ϭ 6.38,
punctella males left the release cage and neither per-
df ϭ 4, P ϭ 0.17), but a signiÞcantly higher proportion
methrin concentration (F4,141 ϭ 1.99; P ϭ 0.10) nor of P. interpunctella males landed on the platform when
ZETA concentration (F1,141 ϭ 3.86; P ϭ 0.06) had the attracticide contained 0.16% ZETA (n ϭ 77, 0.71 Ϯ
signiÞcantly affect on the time the male moths stayed
0.05) compared with 0.32% ZETA (n ϭ 74, 0.55 Ϯ 0.06)
at the release platform before take-off. SigniÞcantly
(2 ϭ 4.16, df ϭ 1, P ϭ 0.04) (Table 1). Flight time of
Fig. 2. Status of individual P. interpunctella males 24 h after exposure to 0.16 or 0.32% ZETA and 0, 3, 6, 12, and 18%
permethrin. The status of the male was assessed according to the following scale: 0, dead; 1, immobile, and/or affected; and
2, alive and not affected (normal behavior). Results with different ZETA concentrations were grouped because only
permethrin concentration had signiÞcant effect on male status (as indicated by different letters).
NANSEN AND PHILLIPS: ATTRACTICIDE FOR P. interpunctellaMean (SE) behavioral responses of P. interpunctella males in wind tunnel to attracticides with different pheromone and permethrin concentrations a Fanning during preconditioning was scored from 0 (no fanning) to 3 (intensive fanning). b Time in seconds before the male left the release cage. c Proportion of males performing upwind ßight. d Proportion of ßying males that arrived at the platform holding the attracticide. e Total ßight time in seconds. f Score for touch with the attracticide: 0, no touch; 1, gentle touch with either a single leg or with just one antenna; 2, touch with at least
two body parts (e.g., two legs, one leg and an antenna) or touch with a wing; and 3, substantial or prolonged contact with the gel.
those landing on the attracticide platform did not vary
ments for moth densities of three and Þve mating
signiÞcantly with neither permethrin concentration
pairs per storage shed (P Ͼ 0.05).
(F4,85 ϭ 0.71; P ϭ 0.59) nor ZETA concentration (F1,85
ϭ 1.27; P ϭ 0.26). The scoring of the maleÕs contact
with the attracticide revealed no signiÞcant effect of
Discussion
permethrin concentration (F4,85 ϭ 0.79; P ϭ 0.53) but
The toxicity tests in this study demonstrated that a
was signiÞcantly higher when the attracticide con-
brief and gentle contact with the attracticide contain-
tained 0.16% ZETA (n ϭ 54, 2.07 Ϯ 0.13) compared
ing at least 3% permethrin affects male survivorship
with 0.32% ZETA (n ϭ 41, 1.71 Ϯ 0.13) (2 ϭ 5.07, df ϭ
and immediate ability to perform courtship behavior
and successful copulation. Nevertheless, some mating
Suppression Study. Results from the toxicity test
may have occurred with intoxicated males before their
revealed no signiÞcant difference in toxicity when
deaths because more eggs were laid in these jars than
permethrin concentration was at least 6%, and the
in jars with unmated females. The glass jars used for
wind tunnel experiment indicated stronger and more
these experiments represent highly constrained envi-
complete anemotactic ßight response to the attracti-
ronments, in which the time needed to search for a
cides containing 0.16% ZETA compared with those
mating partner was reduced to a minimum due to the
containing 0.32%. Consequently, we decided to use
small search area. Hence, mating could theoretically
the attracticide containing 6% permethrin and 0.16%
occur within a few minutes after P. interpunctella
ZETA for the suppression study in storage sheds.
males had been exposed to the attracticide. Other
When all trials (three moth densities ϫ three treat-
work has documented that P. gossypiella males can
ments ϫ six replications) were included in the anal-
recover and locate sex pheromone sources 2Ð4 d after
ysis, there was no signiÞcant difference in mean num-
a sublethal poisoning with an attracticide (Haynes et
ber of oviposited eggs per P. interpunctella female for
al. 1986), but we found that permethrin concentra-
the three attracticide treatments (F
tions Ն6% killed most of the P. interpunctella males
0.29), but the mean number of eggs oviposited per
within a 24-h period. Our current study did not ex-
female decreased signiÞcantly with increasing moth
amine the longevity of permethrin activity after aging,
and such work is needed before a recommendation on
density (F2,49 ϭ 4.60; P ϭ 0.01). Due to the apparent a speciÞc permethrin concentration can be made for
effect of moth density, we conducted a pairwise one-
an attracticide that may be required to kill insects over
way analysis of variance (ANOVA) to examine treat-
several weeks in a practical application.
ment effect within each of the moth densities sep-
The results here, and those from a similar study
arately, and we found a signiÞcant effect of
(Phillips 1994), indicate that relatively higher doses of
attracticide treatment. At the lowest moth density,
ZETA released from a point source elicit fewer con-
the number of eggs laid per female was signiÞcantly
tacts with the source by responding males compared
lower for the trials with one exposed attracticide gel
with lower doses. Responses of male Lepidoptera to
dot compared with control trial (no attracticide)
synthetic sex pheromones can be affected by, among
(F11 ϭ 10.64; P Ͻ 0.01), whereas there was no sig- other variables, relative concentrations and composi-
niÞcant difference between trials with one screened
tion of the materials released from a point source
attracticide gel dot and control (F11 ϭ 1.95; P Ͻ 0.19) (Carde´ and Baker 1984). Although up to four separate
(Fig. 3). There was no signiÞcant difference in the
sex pheromone components have been identiÞed
number of eggs laid per female among the treat-
from P. interpunctella females (Zhu et al. 1999), we
Fig. 3. Three densities of P. interpunctella (females:males: 1:1, 3:3, and 5:5) were released and oviposition in food dishes
was assessed after 76 h in storage sheds with three different treatments: 1) one 0.015-g freely exposed gel dot of 6% permethrin
and 0.16% ZETA was applied to the center of the south wall in the storage shed (dot exposed); 2) one screened gel dot of
was applied to the center of the south wall in the storage shed (screened dot); and 3) control (no dot). Different letters among
treatment indicate signiÞcant difference in number of eggs oviposited in food dishes at the lowest moth density.
used only the predominant compound, ZETA. P. in-
moths at high densities with attracticide may have
terpunctella males respond with a more complete se-
only limited effect on the total population because
quence of orientation and mating behaviors to blends
remaining males tend to compensate by increasing
of pheromone compounds compared with ZETA
their number of matings (Brower 1975). We found
alone (Vick et al. 1981, Zhu et al. 1999). Thus, it is
that when the moth density is one moth pair per 11.3
possible that a higher efÞcacy of a P. interpunctella
m3, signiÞcantly fewer eggs were laid in the simulated
attracticide could be obtained in future work by using
storage sheds with a freely exposed attracticide gel dot
the four-component pheromone blend instead of
compared with the oviposition in storage sheds with a
ZETA alone. Furthermore, if ZETA only were to be
caged dot of attracticide or control (untreated) stor-
used in practice for an attracticide, the formulation
age sheds. Because one attracticide gel dot per 11.3 m3
that would deliver an optimal release rate over a given
deployed under a moth-proof screen cage did not
time period for service would need to be determined.
affect oviposition compared with untreated metal
We showed that the ßight response of P. interpunc-
buildings, this study suggests that mating disruption
tella males to the attracticide was unaffected by a
was not occurring at this amount of the major pher-
permethrin concentration up to18%. Based on recom-
omone component we used in the formulation and this
mendations from the manufacturer of the attracticide
single dot density per 11.3 m3. Rather, the results
gel (IPM Tehnologies Inc.), it was not recommend-
able to increase the permethrin concentration beyond
implicate the toxic effect of the permethrin acting
18% because this would substantially affect the phys-
directly on males that contacted the attracticide as the
ical characteristics (e.g., stickyness) of the gel. Haynes
cause of reduced oviposition. The signiÞcant differ-
et al. (1986) found no evidence of reduction in upwind
ence in total oviposition at different moth pair den-
ßight responses and source contact of P. gossypiella
sities in the metal buildings was not easily interpreted,
males when 1 and 10% of either cypermethrin, per-
and it seems unlikely that three petri dishes of 10 g of
methrin, or fenvalerate were added to an attracticide
oil-treated wheat kernels could be a limiting factor.
pheromone source in attracticide studies, but Phelan
Little is known about intraspeciÞc competition of ovi-
and Baker (1987) found that a 1% concentration of
positing P. interpunctella females, but Phillips and
either cypermethrin or permethrin did reduce the
Strand (1994) found that more oviposition by P. in-
ßight responses of female Ameyelois transitella (Lep-
terpunctella occurred on dishes of food contaminated
idoptera: Pyralidae) in similar wind tunnel tests. Fairly
with conspeciÞc larval secretions than on uncontam-
small dots of attracticide were used in our current
inated food. However, it is not known whether eggs
study (15 mg), so it is possible that repellency to
laid in wheat kernels are attractive to ovipositing P.
permethrin would have been detected if larger dots
interpunctella females as well.
This study with small storage sheds provides the
Control tactics based on mass-killing of male insects
baseline for similar controlled studies in larger struc-
will generally be more effective at lower population
tures with various moth densities and attracticide de-
densities (Lanier 1990). Hence, suppression of male
ployment schemes for further evaluation of the at-
NANSEN AND PHILLIPS: ATTRACTICIDE FOR P. interpunctella
tracticide as a suppression tool for P. interpunctella
comparison with the malesÕ response to sex pheromone. Haynes, K. F., W. G. Li, and T. C. Baker. 1986. Control of
pink bollworn moth (Lepidoptera: Gelechidae) with in-
Acknowledgments
secticides and pheromones (attracticide): lethal and sub-
lethal effects. J. Econ Entomol. 79: 1466Ð1471.
We thank Drs. James F. Campbell and Michael D. Toews
Johnson, J. A., P. L. Wofford, and L. C. Whitehand. 1992.
for reviews of earlier drafts of this article. IPM Technologies,
Effect of diet and temperature on development rates,
Inc. was instrumental in formulating and providing the at-
survival, and reproduction of the Indianmeal moth (Lep-
tracticides. We appreciate the technical assistance of Stacy
idoptera: Pyralidae). J. Econ. Entomol. 85: 561Ð566.
Sanders in conducting the experiments. This research was
Kuwahara, Y., and J. E. Casida. 1973. Quantitative analysis of
supported in part by a grant from the USDA, Cooperative and
the sex pheromone of several phycitid moths by electron-
State Research, Education and Extension Service in the Risk
capture gas chromatography. Agric. Biol. Chem. 37: 681Ð
Avoidance and Mitigation Program, agreement no.
00Ð51101Ð9674 and was approved for publication by the
Kuwahara, Y., C. Kitamura, S. Takahashi, H. Hara, S. Ishii,
Oklahoma Agricultural Experiment Station. and H. Fukami. 1971. Sex pheromone of the Almond
moth and the Indian Meal moth: cis-9, trans-12-Tetrade-
cadienyl Acetate. Science 171: 801Ð802. References Cited Lanier, G. N. 1990. Principle of attraction-annihilation: Brady, U. E., J. H. Tumlinson, R. G. Brownlee, and R. M.
mass trapping and other means, pp. 25Ð45. In R. L. Ridg-
Silverstein. 1971. Sex stimulant and attractant in the In-
way, R. M. Silverstein, and M. N. Inscoe, [eds.], Behavior-
dian meal moth and almond moth. Science (Wash. DC)
modifying chemicals for insect management, applications
of pheromones and other attractants. Marcel Dekker,
Brockerhoff, E. G., and D. M. Suckling. 1999. Development
of an attracticide against light brown apple moth (Lep-
Lecato, G. L. 1976. Yield, development, and weight of Cadra
idoptera: Tortricidae). J. Econ. Entomol. 92: 853Ð859. cautella (Walker) and Plodia interpunctella (Hu¨bner) on
Brower, J. H. 1975. Plodia interpunctella: effect of sex ratio
twenty-one diets derived from natural products. J. Stored
on reproductivity. Ann. Entomol. Soc. Am. 68: 847Ð851. Mafra-Neto, A., and T. C. Baker. 1996. Timed, metered ´, R. T., and T. C. Baker. 1984. Sexual communication
with pheromones, pp. 355Ð383. In W. J. Bell and R. T.
sprays of pheromone disrupt mating of Cadra cautella
(Lepidoptera: Pyralidae). J. Agric. Entomol. 13: 149Ð168.
´ [eds.], Chemical ecology of insects. Chapman &
Miller, E., R. T. Staten, C. Nowell, and J. Gourd. 1990. Pink
bollworm (Lepidoptera: Gelechiidae): point source den-
´, R. T., and A. K. Minks. 1995. Control of moth pests
by mating disruption: successes and constraints. Annu.
sity and its relationship to efÞcacy in attracticide formu-
lations of gossyplure. J. Econ. Entomol. 83: 1321Ð1325. Charmillot, P. J., and D. Hofer. 1997. Control of codling Nansen, C., and T. W. Phillips. 2003. Ovipositional re-
moth, Cydia pomonella, by an attract and kill formulation.
sponses of the Indianmeal moth, Plodia interpunctella
Technical transfer in mating disruption. Int. Org. Biocon-
(Hu¨bner) (Lepidoptera: Pyralidae) to oils. Ann. Ento-
Cline, L. D. 1978. Penetration of seven common ßexible Nansen, C., T. W. Phillips, M. N. Parajulee, and R. A. Franqui-
packing materials by larvae and adults of eleven species
Rivera. 2004. Comparison of direct and indirect sam-
of stored-product insects. J. Econ. Entomol. 71: 726Ð729.
pling procedures Plodia interpunctella in a corn storage
Cline, L. D., and H. A. Highland. 1985. Survival, reproduc-
facility. J. Stored Prod. Res. 40: 151Ð168.
tion, and development of seven species of stored-product
Phelan, P. L., and T. C. Baker. 1987. An attracticide for
insects on the various food components of lightweight,
control of Amyelois transitella (Lepidoptera: Pyralidae)
high-density, prototype military rations. J. Econ. Ento-
in almonds. J. Econ. Entomol. 80: 779Ð783. Phillips, T. W. 1994. Pheromone of stored-product insects: Cuperus, G. W., R. T. Noyes, W. S. Fargo, B. L. Clary, D. C.
current status and future perspectives. pp. 479Ð486. In E. Arnold, and K. Anderson. 1990. Management practices
Highley, E. J. Wright, H. J. Banks, and B. R. Champ [eds.],
in a high-risk stored-wheat system in Oklahoma. Am.
Proceedigns fo the 6th International Working Confer-
ence on Stored-Product Protection, 17Ð23 April 1994,
De Souza, K. R., L. J. Mcveigh, and D. J. Wright. 1992.
Selection of insecticides for lure and kill studies against
Phillips, T. W. 1997. Semiochemicals of stored-product in- Spodoptera littoralis (Lepidoptera: Noctuidae). J. Econ.
sects: Research and applications. J. Stored Prod. Res. 33:
Doud, C. W., and T. W. Phillips. 2000. Activity of Plodia Phillips, T. W., and M. R. Strand. 1994. Larval secretions interpunctella (Lepidoptera: Pyralidae) in and around
and food odors affect orientation in female Plodia inter-
ßour mills. J. Econ. Entomol. 93: 1842Ð1847. punctella. Entomol. Exp. Appl. 71: 185Ð192. Downham, M.C.A., L. J. Mcveigh, and G. M. Moawad. 1995. Phillips, T. W., R. C. Berberet, and G. W. Cuperus. 2000a.
Field investigation of an attracticide control technique
Post-harvest integrated pest management, pp. 2690Ð2701.
using the sex pheromone of the Egyptian cotton leaf-
In F. J. Francis [ed.], Encyclopedia of food science and
worm, Spodoptera littoralis (Lepidoptera: Noctuidae).
technology, 2nd ed. Wiley, New York. Phillips, T. W., P. M. Cogan, and H. Y. Fadamiro. 2000b. Fields, P. G., and N.D.G. White. 2002. Alternative to methyl
Pheromones, pp. 273Ð302. In B. Subramanyam and D. W.
bromide treatments for stored-product and quarantine
Hagstrum [eds.], Alternatives to pesticides in stored-
insects. Annu. Rev. Entomol. 47: 331Ð359.
product IPM. Kluwer Academic Publishers. Boston. Haynes, K. F., and T. C. Baker. 1989. An analysis of anemo- Ryne, C., G. P. Svensson, and C. Lo¨fstedt. 2001. Mating dis-
tactic ßight in female moths stimulated by host odour and
ruption of Plodia interpunctella in small-scale plots: ef-
fects of pheromone blend, emission rates, and population
the control of Ephestia kuehniella Zeller: studies of ef-
density. J. Chem. Ecol. 27: 2109Ð2124.
fectiveness. J. Appl. Entomol. 111: 451Ð456. SAS Institute. 1999. SAS/STAT userÕs guide, version 8. SAS Storey, C. L., D. B. Sauer, and D. Walker. 1983. Insect pop-
ulations in wheat, corn, and oats stored on the farm. J. Shani, A., and J. Clearwater. 2001. Evasion of mating dis-
ruption in Ephestia cautella (Walker) by increased pher-
Vick, K. W., J. A. Coffelt, and M. A. Sullivan. 1978. Disrup-
omone production relative to that of undisrupted popu-
tion of pheromone communication in the angoumois
lations. J. Stored Prod. Res. 37: 237Ð252.
grain moth with synthetic female sex pheromone. Envi-
Soderstrom, E. L., D. G. Brandl, K. W. Vick, and J. A. Coffelt. 1980. Evaluation of synthetic sex pheromone. Insecticide Vick, K. W., J. A. Coffelt, R. W. Mankin, and E. L. Soder- stroem. 1981. Recent developments in the use of pher- Sower, L. L., and M. D. Shorb. 1985. Pesticides have little
omones to monitor Plodia interpunctella and Ephestia
impact on attraction of three species of male moths to sex
cautella, pp. 19Ð28. In E. R. Mitchell (ed.), Management
pheromone. J. Econ. Entomol. 78: 908Ð912.
of insect pests with semiochemicals. Plenum Publ., New
Sower, L. L., K. W. Vick, and J. H Tumlinson. 1974. (Z,E)-
9,12-Tetradecadien-1-ol: a chemical released by female Plo-Vick, K. W., P. G. Koehler, and J. J. Neal. 1986. Incidence of dia interpunctella that inhibits the sex pheromone response
stored-product Phycitinae moth in food distribution
of male Cadra cautella. Environ. Entomol. 3: 120Ð122.
warehouses as determined by sex pheromone-baited
Suckling, D. M., and E. G. Brockerhoff. 1999. Control of
traps. J. Econ. Entomol. 79: 936Ð939.
light brown apple moth (Lepidoptera: Tortricidae) using
Zhu, J., C. Ryne, R. Unelius, P. G. Valeur, and C. Lo¨fstedt.
an attracticide. J. Econ. Entomol. 92: 367Ð372. 1999. ReidentiÞcation of the female sex pheromone of Teal, P.E.A., R. R. Heath, B. D. Dueben, J. A. Coffelt, and
the Indian meal moth, P. interpunctella: evidence for a
K. W. Vick. 1995. Production and release of (Z,E)-9,12-
four-component pheromone blend. Entomol. Exp. Appl.
Tetradecadienal by sex pheromone glands of female Plo-dia interpunctella (Lepidoptera: Pyralidae). J. Chem. Received for publication 13 August 2003; accepted 20 Oc-Trematerra, P., and A. Capizzi. 1991. Attracticide method in
Referencing Guidelines A citation is a passage or opinion from a book, article or webpage etc., made in such a way that a reader can identify it. A reference is a direction to a book, article or webpage etc., where certain information may be found; an indication of the author, work, page etc., to be looked at or consulted. A reference list is a collection of references. Referenci
Guidelines for the Management of Asthma: 2007 Update Jennifer Lee, PharmD, BCPS The University of Connecticut, School of Pharmacy is accredited by the Accreditation Council for Pharmacy Education as a provider of continuing pharmacy education. This program Guidelines for the Management of Asthma: 2007 Update (ACPE No. 009-000-08-023-H01-P), is approved for 0.10 CEU or 1 Contact Hour of contin
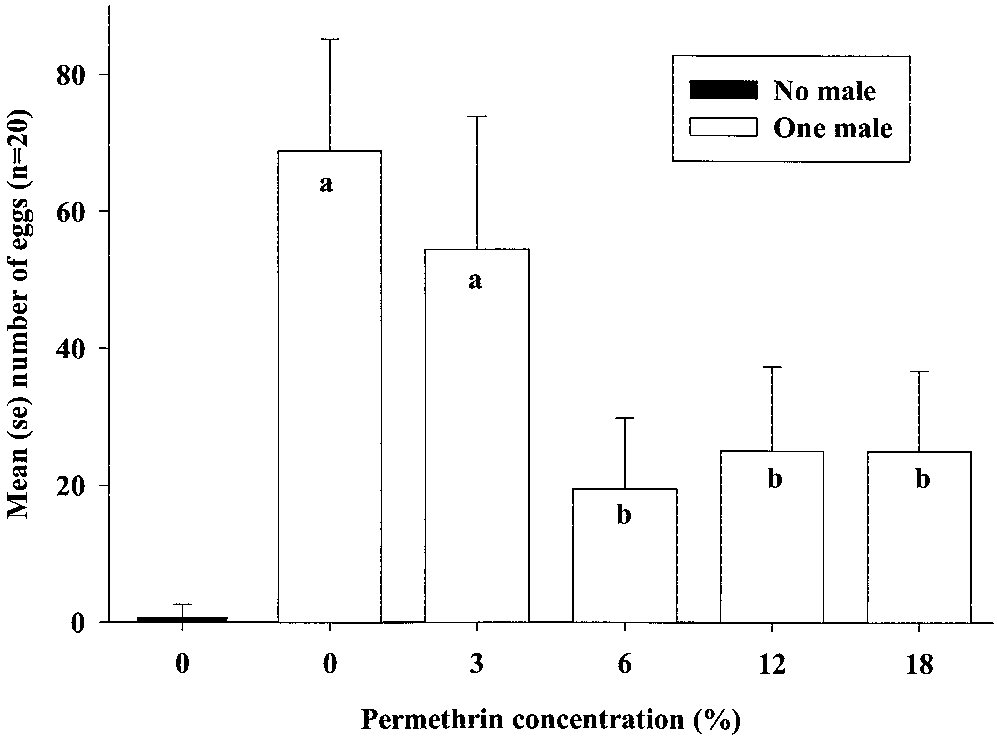
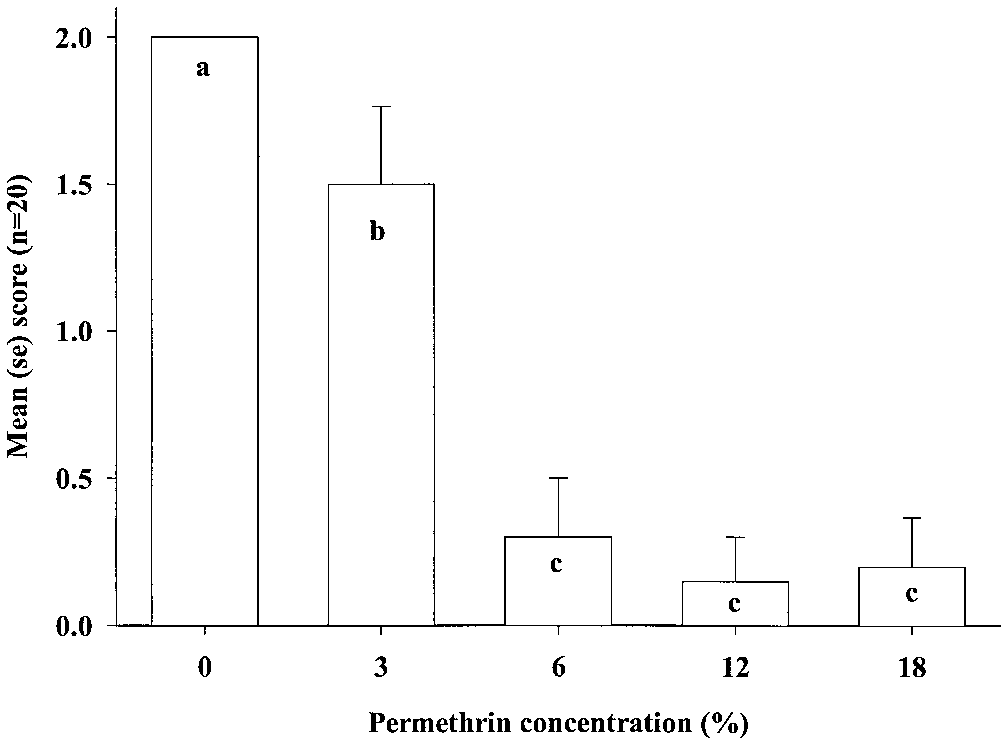 Fig. 1. Mean oviposition within 24 h of single P. interpunctella couples after the male had subtle contact with one of 10
Fig. 1. Mean oviposition within 24 h of single P. interpunctella couples after the male had subtle contact with one of 10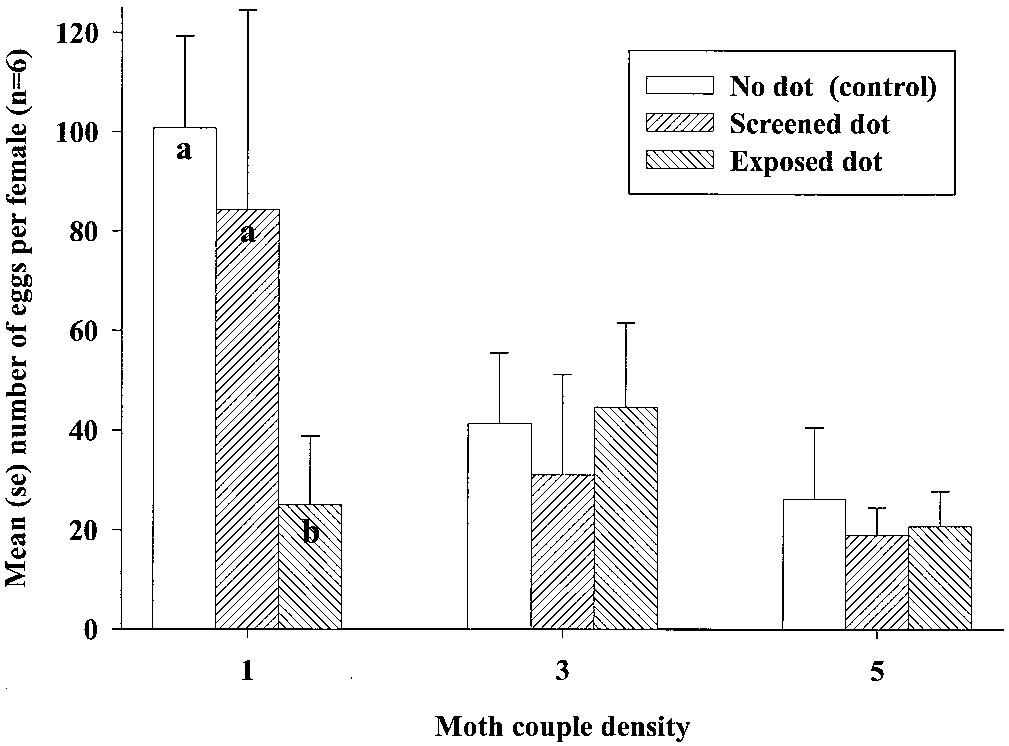 Fig. 3. Three densities of P. interpunctella (females:males: 1:1, 3:3, and 5:5) were released and oviposition in food dishes
Fig. 3. Three densities of P. interpunctella (females:males: 1:1, 3:3, and 5:5) were released and oviposition in food dishes