Le tadalafil possède une affinité marquée pour la PDE5, mais épargne en grande partie les isoformes PDE1, PDE2 et PDE11, réduisant ainsi le risque d’effets extra-caverneux. L’action se traduit par une augmentation contrôlée de la circulation sanguine locale, indépendante des variations alimentaires. Sa pharmacocinétique repose sur une absorption digestive rapide, un métabolisme hépatique par CYP3A4 et une distribution tissulaire large. La biodisponibilité reste stable, et l’équilibre plasmatique est atteint en quelques jours lors d’administrations répétées. Les interactions cliniquement significatives surviennent avec les inhibiteurs puissants de CYP3A4 tels que le kétoconazole. Dans la littérature pharmacologique, acheter cialis 20 mg est souvent associé à des schémas d’utilisation basés sur la durée prolongée de son action.
Antimicrobianos.com.ar
ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, May 2002, p. 1462–1468
0066-4804/02/$04.00ϩ0 DOI: 10.1128/AAC.46.5.1462–1468.2002
Copyright 2002, American Society for Microbiology. All Rights Reserved.
Plasmidic Extended-Spectrum -Lactamases in Vibrio cholerae O1
Alejandro Petroni,1 Alejandra Corso,1 Roberto Melano,1 Marı´a Luisa Cacace,2 Ana Marı´a Bru,3
Servicio Antimicrobianos, Instituto Nacional de Enfermedades Infecciosas, ANLIS “Dr. Carlos G. Malbrán,” Buenos Aires,1 andHospital San Vicente de Paul, Orán,2 and Hospital Juan D. Perón, Tartagal,3 Salta, Argentina
Received 6 June 2001/Returned for modification 29 September 2001/Accepted 18 January 2002
Since 1992 there have been seven major outbreaks of cholera in Argentina. Susceptibility analysis of 1,947 isolates (40% of reported cases) of Vibrio cholerae O1 biotype El Tor suggested the presence of extended- spectrum -lactamases (ESBLs) in 28 isolates. Because of their different susceptibility profiles, V. cholerae isolates M1502, M1516, M1573, and M3030 (all of which are of the Ogawa serotype) were selected for the present study. By susceptibility analysis, isoelectric focusing, and PCR-based restriction fragment length polymorphism analysis, CTX-M-type enzymes were identified in three isolates, whereas a PER-2-type enzyme, in addition to a TEM-1-like enzyme, was identified in the other isolate. The presence of these ESBLs in V. cholerae isolates resulted in MICs well below those commonly observed for members of the family Enterobac- teriaceae. Genes that encode both ESBLs were transferred to Escherichia coli by conjugation, together with all determinants of resistance to non--lactam antibiotics (gentamicin, kanamycin, and sulfamethoxazole for all isolates; amikacin and streptomycin for three isolates; trimethoprim, tetracycline, and chloramphenicol for two isolates). Plasmid profile analysis and Southern blotting revealed the presence of single plasmids of about 150 kb in the four V. cholerae isolates and their respective transconjugants and revealed that the plasmids harbored genes encoding CTX-M-type or PER-2-type ESBLs. These results strongly suggest the broad spread of these ESBLs among genera belong to families other than the Enterobacteriaceae.
Extended-spectrum -lactamases (ESBLs) have been classi-
Pasteran, R. G. Melano, M. F. Galas, M. M. Rodriguez, WHO-
fied by Bush et al. (8) in functional group 2be or in Ambler
NET-Argentina Collaborative Group, and M. A. Rossi, 39th
molecular class A. These enzymes include in their spectra of
Intersci. Conf. Antimicrob. Agents Chemother., abstr. 1475,
activity penicillins, most cephalosporins, and aztreonam; they
1999). Second, there are the CTX-M -lactamases, which are
do not hydrolyze cephamycins (cefoxitin, cefotetan, and moxa-
more active against cefotaxime than ceftazidime (for a recent
lactam) and carbapenems (imipenem and meropenem); and
review, see reference 45). These enzymes have been observed
they are inhibited by clavulanic acid, tazobactam, and sulbac-
in members of the family Enterobacteriaceae since 1990, and 14
tam (21). Since 1983, plasmidic ESBLs have been extensively
members of the family have been reported to date (5, 6, 12, 18,
reported worldwide, mainly among species of the family En-terobacteriaceae (17, 21; see http://www.lahey.org/studies/webt
Nowadays, CTX-M-producing enterobacteria appear to be
.htm). Most plasmidic ESBLs belong to the TEM or SHV type
widely spread over very distant geographic regions including
(21). Two novel sets of ESBLs that are unrelated to the TEM
Japan and Taiwan (45, 49), several European countries (9, 12,
and SHV types and that belong to group 2be of Bush et al. (8)
27, 36, 38, 45), South America (5, 6, 45), and very recently,
have been observed. First, there are the PER-type enzymes (3,
Kenya (18). In particular, CTX-M-2 was broadly detected
25, 26), which are more active against ceftazidime than cefo-
mainly in Argentina and also in neighboring countries (31, 35,
taxime. PER-1 was isolated from Pseudomonas aeruginosa
45; Galas et al., 38th ICAAC)). Comparisons of the amino acid
(26), Salmonella enterica serovar Typhimurium (46), and Aci-
sequences of the CTX-M family showed four clusters or
netobacter spp. (47), whereas PER-2 was reported in members
groups. The first group includes CTX-M-1, CTX-M-3, CTX-
of the family Enterobacteriaceae from Argentina (3; M. F. Ga-
M-10, and CTX-M-12; the second group comprises CTX-M-2,
las, F. G. Pasterán, R. G. Melano, A. E. Petroni, G. Lopez,
A. C. Corso, M. A. Rossi, and WHONET Collaborative
CTX-M-4 to CTX-M-7, and Toho-1; the third group contains
Group, 38th Intersci. Conf. Antimicrob. Agents Chemother.,
Toho-2, CTX-M-9, and CTX-M-16; and the fourth group con-
abstr. E109, 1998; M. F. Galas, M. J. Rapoport, F. G. Pasterán,
R. G. Melano, A. E. Petroni, P. G. Ceriana, WHONET-Ar-
The broad spread of these enzymes among human patho-
gentina Collaborative Group, and M. A. Rossi, 39th Intersci.
genic bacteria which have never been shown to harbor ESBLs
Conf. Antimicrob. Agents Chemother., abstr. 1474, 1999; F. G.
could constitute a major health concern and an epidemiologicproblem. Since 1991 Vibrio cholerae O1 biotype El Tor hasbecome one of the most clinically important reemerging patho-
* Corresponding author. Mailing address: Servicio Antimicrobianos,
gens in Latin America because it has caused cholera outbreaks
INEI-ANLIS “Dr. Carlos G. Malbrán,” Av. Velez Sarsfield 563 (1281),
and endemic diarrhea. From 1992 to 1998, during seven chol-
Buenos Aires, Argentina. Phone and Fax: 54-11-4303-2812. E-mail:
era seasons in Argentina, 1,947 isolates of V. cholerae O1
† Dedicated to the memory of Alicia Rossi.
biotype El Tor (40.3% of isolates from reported cases) were
EXTENDED-SPECTRUM -LACTAMASES IN V. CHOLERAE
TABLE 1. Susceptibility profile distributions for ESC-resistant V. cholerae isolates from the seven Argentinean cholera seasons
ESC-resistant isolates (no. of isolates)da Cholera seasons comprised time periods which generally extended from October to May. b Reported cases were recorded by the Sistema Nacional de Vigilancia Epidemiológica, Ministerio Nacional de Salud y Acción Social. c Number of isolates analyzed at the National Reference Laboratory (NRL). d Resistance or reduced susceptibility are indicated. Abbreviations: AMK, amikacin; AMP, ampicillin; ATM, aztreonam; CAZ, ceftazidime; CFP, cefoperazone;
CHL, chloramphenicol; CTX, cefotaxime; ERY; erythromycin; GEN, gentamicin; KAN, kanamycin; NIT, nitrofurantoin; O129, vibriostatic agent; SFI, sulfisoxazole;
STR, streptomycin; SXT; trimethoprim-sulfamethoxazole; TET, tetracycline. e ND, no ESC-resistant isolates were detected.
received at the National Reference Laboratory of Argentina.
in 24 cases in seven families. The representative isolate selected for this study
Ampicillin resistance was detected in 34 of 1,947 isolates. Sur-
(isolate M3030) was isolated from a 48-year-old woman in the first family af-
prisingly, 28 of the ampicillin-resistant isolates showed strong
fected; she had initially been treated with a single dose of doxycycline.
A susceptible V. cholerae 2717 O1 biotype El Tor isolate used in the suscep-
reductions in their susceptibilities to extended-spectrum ceph-
tibility profile analysis was isolated (February 1993) from a 16-month-old female
alosporins (ESCs) (cefotaxime and/or ceftazidime), cefopera-
patient with diarrhea, vomiting, and dehydration symptoms.
zone, and aztreonam. ESC resistance was reversed by clavu-
Spontaneous mutants of Escherichia coli C600 and ER1793 (New England
lanic acid, suggesting the presence of ESBLs. In this work, we
Biolabs, Beverly, Mass.) resistant to nalidixic acid and rifampin were obtained as
report on the first identification in V. cholerae of two plasmid-
described previously (37) and were used as recipient strains for bacterial conju-
mediated ESBLs: CTX-M-type and PER-2-type ESBLs. These
Media and chemicals. Mueller-Hinton broth and agar, tryptic soy broth, and
facts strongly suggest the broad spread of these enzymes
Luria-Bertani (LB) broth and agar were from Difco (Detroit, Mich.). Restriction
among genera belonging to families other than the Enterobac-
enzymes were from New England Biolabs. Routine chemicals were from Merck
(This study was presented in part at the 38th Interscience
Antimicrobial agents and susceptibility testing. The following antimicro-
Conference on Antimicrobial Agents and Chemotherapy, 24 to
bial agents were obtained from standard laboratory powders and the indicatedsources: amikacin, aztreonam, and cefepime, Bristol-Myers Squibb; amoxicillin,
27 September 1998, San Diego, Calif. [M. Galas, A. Petroni, R.
Roemmers; clavulanic acid, SmithKline Beecham; ampicillin, Temis Lostaló;
Melano, A. Corso, M. Rodriguez, M. L. Cacace, A. M. Bru,
chloramphenicol, Parke-Davis; cefotaxime, Argentia; cefoxitin, imipenem, and
and A. Rossi, Abstr. 38th Intersci. Conf. Antimicrob. Agents
norfloxacin, Merck Sharp & Dohme; ceftazidime, Glaxo; ceftibuten, gentamicin,
nitrofurantoin, sulfamethoxazole, and trimethoprim, Schering Plough; cephalo-thin, erythromycin, piperacillin, streptomycin, and tazobactam, Wyeth-Ayerst;kanamycin, Armstrong; sulfisoxazole, Abbott; tetracycline, Microsules y Bern-
MATERIALS AND METHODS
abó; and vibriostatic agent O129, Sigma.
MICs were determined by the agar dilution technique according to the spec-
Bacterial strains. From February 1992 to June 1998, a total of 1,947 V. cholerae isolates were received at the National Reference Laboratory (Table 1).
ifications of NCCLS (23) by adoption of the criteria for the family Enterobacte-
Susceptibility analysis was carried out by the disk diffusion method according to
riaceae for antimicrobial agents other than ampicillin, tetracycline, trimethoprim-
the specifications of NCCLS (24) (adopting the criteria for the family Enterobac-
sulfamethoxazole, chloramphenicol, and sulfonamides. Clavulanic acid was used
teriaceae for antimicrobial agents other than ampicillin, tetracycline, tri-
at a ratio of 1 to 2 in combination with amoxicillin or at a constant concentration
methoprim-sulfamethoxazole, chloramphenicol, and sulfonamides), and 28 iso-
of 4 g/ml in combinations with other -lactams. Tazobactam was used at a
lates showed resistance to cefotaxime and/or ceftazidime, cefoperazone, and
constant concentration of 4 g/ml.
aztreonam. These V. cholerae isolates (all isolates were of the Ogawa serotype)
The following reference strains were used for quality control of dilution tests:
were grouped according to their susceptibility profiles, and one isolate from each
P. aeruginosa ATCC 27853, Staphylococcus aureus ATCC 29213, and E. coli
group was selected to carry out this study, namely, isolates M1502, M1516,
ATCC 25922 and ATCC 35218. Since the MICs of antimicrobial agents for V.
M1573, and M3030 (Table 1). V. cholerae M1502 has already been described
cholerae 2717 were in the range of those previously reported for susceptible
(34). V. cholerae M1516 was isolated from a 43-year-old female outpatient with
isolates (34, 48), this isolate was used as a control strain for analysis of the V.
vomiting and moderate dehydration in Tartagal, Argentina (north of Salta Prov-
cholerae O1 biotype El Tor susceptibility profile.
ince), in March 1993. This isolate was one of three isolates that had identical
Conjugation assays. Biparental conjugations were performed as follows. Cells
susceptibility profiles and that had been detected in members of the same family.
of both the donor and the recipient strains were mixed on LB agar at a ratio of
V. cholerae M1573 was isolated from a hospitalized newborn male with diarrhea
5 to 1, and the mixture was incubated for 18 h at 35°C. Transconjugants were
who had been treated with ampicillin plus gentamicin in Tartagal (April 1993).
selected on Mueller-Hinton agar supplemented with nalidixic acid (50 g/ml)
ESC-resistant V. cholerae isolates were not detected during the following three
and cefotaxime (8 g/ml) or ceftazidime (0.5 g/ml), as indicated.
cholera seasons (a 4-year period). Toward the end of the sixth cholera season
-Lactamase preparation. Bacterial cultures were grown overnight at 35°C in
(March to April 1997) a small outbreak due to an ESC-resistant strain of V.
10 ml of brain heart infusion broth. Cells were harvested by centrifugation,
cholerae O1 occurred in Irigoyen and Pichanal, Argentina (north of Salta Prov-
washed, and resuspended in 0.5 ml of 10 mM phosphate buffer (pH 7.0). Cell
ince). The outbreak began with a 1-year-old male patient who acquired the
suspensions were disrupted at 4°C by sonication (two pulses of 45 s at 20 Hz,
illness during a hospitalization and rapidly spread into the community, resulting
separated by an interval of 30 s, on a Vibra Cell Sonifier [Sonics and Materials
Inc., Danbury, Conn.]) and centrifuged (17,000 ϫ g for 10 min at 4°C) for debris
buten, and aztreonam were 64, 64, and 16 times higher, re-
removal. The supernatants were used for isoelectric focusing (IEF) assays.
spectively, for the group B isolate than for the group A isolates. IEF of -lactamases. Crude -lactamase extracts were focused on broad-range
In order to analyze if the V. cholerae resistance determinants
precast polyacrylamide gels (pH 3.5 to 9.5; Ampholine PAGplate; PharmaciaBiotech, Uppsala, Sweden) with a Multiphor II apparatus (Pharmacia LKB,
were harbored in conjugative plasmids, isolates M1502 and
Uppsala, Sweden), according to the instructions of the manufacturer. -Lacta-
M1573 were mated with E. coli C600 (selections were carried
mase bands were visualized by the iodometric method described by Labia and
out with nalidixic acid plus cefotaxime for M1502 or with
Barthélémy (20), with minor modifications. To differentiate between bands pro-
nalidixic acid plus ceftazidime for M1573), rendering transcon-
duced by broad-spectrum -lactamase or ESBL activities, after IEF the gel wascovered with an agar overlay containing cephaloridine (1 mg/ml) plus penicillin
jugants M1538 and M1561, respectively. In addition, V. chol-
(0.1 mg/ml) or ceftazidime (1 mg/ml) plus ceftriaxone (1 mg/ml) as developing
erae M1516 and M3030 were conjugated with E. coli ER1793
substrates, respectively. The gel was incubated at room temperature until the
(selections were achieved with nalidixic acid plus cefotaxime),
bands could be visualized. For -lactamase inhibition procedures, the gel was
rendering transconjugants M3099 and M3033, respectively. The
covered with a piece of filter paper impregnated with 1 mM clavulanic acid or
susceptibility profile of each E. coli transconjugant matched
cefoxitin, as indicated, and incubated for 10 min at room temperature beforedevelopment, as described above. The following -lactamases with known pIs
that of the respective donor V. cholerae isolate (Table 2). For
were used as standards: TEM-1 (pI 5.4), PER-2 (pI 5.4), SHV-2 (pI 7.6), P99 (pI
the group A transconjugant strains, the MICs of cefotaxime
7.8), CTX-M-2 (pI 7.9), and SHV-5 (pI 8.2).
(64 to 128 g/ml) and cefepime (16 to 64 g/ml) were the
PCR amplifications and PCR-RFLP analysis. Primers specific for the TEM
highest and the MICs of ceftazidime (2 to 8 g/ml), ceftibuten
and SHV -lactamase gene families (primers PTEM and PSHV, respectively) (1)
were used for PCRs. For amplification of complete structural genes highly
(0.5 to 1 g/ml), and aztreonam (16 to 64 g/ml) were the
related to that for the CTX-M-2 enzyme (bla
lowest. Otherwise, for the group B transconjugant strain, the
were designed from its reported DNA sequence (2): 5Ј-CGGAATTCATGATG
MICs of ceftazidime (128 g/ml), ceftibuten (32 g/ml), and
ACTCAGAGCATTCG-3Ј and 5Ј-GCTCTAGATTATTGCATCAGAAACCG
aztreonam (128 g/ml) were the highest and the MICs of
TG-3Ј (primers PCTX-M-2 forward and PCTX-M-2 reverse, respectively). For the
cefotaxime (8 g/ml) and cefepime (8 g/ml) were the lowest.
identification of the gene encoding PER-2-type enzymes (blaPER-2-type), primers
5Ј-GTAGTATCAGCCCAATCCCC-3Ј and 5Ј-CCAATAAAGGCCGTCCATC
For both groups A and B, for either V. cholerae isolates or
E. coli transconjugants, clavulanate reduced the MICs nearly
PER forward and PPER reverse, respectively) were designed from
the reported DNA sequence (3). PCR amplifications were performed with a
to the MICs for the susceptible strains.
Perkin-Elmer Cetus thermal cycler in a final volume of 50 l containing 20 pmol
Even though selection of transconjugants was achieved only
of each primer, 25 M each deoxynucleoside triphosphate, 1.5 mM MgCl2, and
2.5 U of Taq polymerase (Promega, Madison, Wis.). DNA templates were
with cefotaxime or ceftazidime, determinants of resistance to
prepared by lysing one or two colonies of V. cholerae isolates or E. coli transcon-
all non--lactam antibiotics, with the exception of reduced
jugants in 50 l of boiling water, as described previously (28, 29). Two microliters
susceptibility to nitrofurantoin, shown by V. cholerae M3030,
was used for the PCR. The PCR program was 10 min of denaturation at 94°C;
were also transferred to E. coli: gentamicin, kanamycin, and
25 cycles of 30 s of denaturation at 94°C, 30 s of annealing at 55°C, and 30 s of
sulfisoxazole for four isolates; amikacin and streptomycin for
extension at 72°C; and a final extension step of 5 min at 72°C. The resultingamplification products were run in a 1.5% agarose gel and purified by using a
three isolates; and trimethoprim, tetracycline, and chloram-
Wizard PCR Preps DNA Purification System (Promega), according to the rec-
phenicol for M1516 and M3030, which were also resistant to
ommendations of the manufacturer. Purified products were used for PCR-based
restriction fragment length polymorphism (PCR-RFLP) analysis, as described
IEF of -lactamases. IEF of crude homogenates of group A
previously (37), or for construction of the labeled probes used in the hybridiza-tion assays. V. cholerae isolates (M1502, M1516, and M3030) and their
Plasmid profile analysis and hybridization. Plasmid DNA was purified from
respective E. coli transconjugants (M1538, M3099, and M3033)
V. cholerae or E. coli transconjugant cells as described previously (22). Plasmidic
showed both broad-spectrum -lactamase activities at pI 5.4
DNAs (about 250 ng) were analyzed by electrophoresis in 0.7% agarose gels
and ESBL activities at the same pI as that for the CTX-M-2
(Tris-acetate buffer), by using plasmids of known size as standards, and were then
standard (pI 7.9). All these activities were inhibited by clavu-
transferred onto and immobilized on a nylon transfer membrane (Zeta-ProbeGT; Bio-Rad, Richmond, Calif.). DNA-DNA hybridizations were performed by
lanic acid. IEF of crude extracts of V. cholerae M1573 and
the method described by Southern (39). Labeling of purified PCR products used
E. coli M1561 (group B) revealed the presence of ESBL activ-
as probes and stripping and reprobing of the membranes were performed with a
ities at the same pI as that for the PER-2 standard (pI 5.4), and
Dig DNA Labeling and Detection kit (Boehringer Mannheim, Mannheim, Ger-
the activities were also susceptible to clavulanic acid inhibition.
many) by the procedures recommended by the manufacturer.
The presence of a broad-spectrum -lactamase activity at pI
5.4 in M1573 and M1561 could not be investigated by the
standard IEF assay because of the overlapping ESBL band at
Antibiotic susceptibilities of V. cholerae isolates and conju-
the same pI. Therefore, in situ inhibition with cefoxitin was
gational transfer of resistance. The susceptibility profiles of V.
performed on the IEF gel. The ESBL activity at pI 5.4, like the
cholerae M1502, M1516, M1573, and M3030 are given in Table
activity of the PER-2 standard, was inhibited, whereas the
2. With the exception of the MICs of cefoxitin and imipenem,
broad-spectrum -lactamase activity at pI 5.4 and the ESBL
the MICs of all -lactams tested were higher for these four
isolates than for susceptible isolate 2717. Two different suscep-
Identification of -lactamases by PCR-RFLP analysis. Both
tibility profiles for -lactams were recognized, and the four
-lactam susceptibility profiles and IEF analysis suggested the
isolates were grouped into groups A (V. cholerae M1502,
presence of a CTX-M-type enzyme and a PER-like enzyme in
M1516, and M3030) and B (isolate M1573). The MICs of
the transconjugant strains of groups A and B, respectively.
cephalothin, cefotaxime, and cefepime were eight, four, and
Therefore, PCR assays were carried out with total DNA from
two times higher, respectively, for the group A isolates than for
the four V. cholerae isolates and their respective E. coli
the group B isolate, whereas the MICs of ceftazidime and
transconjugants as templates and with primers PCTX-M-2, PPER,
ceftibuten for the group A isolates were essentially the same as
PTEM, or PSHV. When primers PCTX-M-2 were used, only tem-
those for V. cholerae 2717. The MICs of ceftazidime, cefti-
plates from the group A V. cholerae isolates and their respec-
EXTENDED-SPECTRUM -LACTAMASES IN V. CHOLERAE
TABLE 2. Antibiotic susceptibilities of V. cholerae isolates and E. coli transconjugants
a Abbreviations: AMC, amoxicillin-clavulanic acid; CLA, clavulanic acid; CTB, ceftibuten; CEF, cephalothin; FOX, cefoxitin; FEP, cefepime; IPM, imipenem; NOR,
norfloxacin; PIP, piperacillin; TZP, piperacillin-tazobactam; TMP; trimethoprim. The abbreviations for the remaining agents are given in footnote d of Table 1. b V. cholerae 2717 is a susceptible isolate. E. coli C600 and ER1793 (STRr) were used as recipient strains for bacterial conjugations. c First-line antibiotics for treatment of V. cholerae infections (4). d The use of nitrofurantoin is equivalent to the use of furazolidone, the MICs of which for susceptible isolates from Argentina were in the range 0.5 to 0.12 g/ml.
tive transconjugants produced amplification products which
To differentiate among these possibilities, we designed a PCR-
were of the same size (900 bp), as expected from the compu-
RFLP analysis with restriction enzyme BsaHI, which includes
tational analysis of the reported DNA sequences of the genes
in its recognition sequence the single point mutation between
highly related to blaCTX-M-2. The group of highly related genes
the CTX-M-2 and the Toho-1 structural genes (Table 3). The
comprised genes encoding CTX-M-2 (2), CTX-M-4 (13),
amplification products obtained were not digested by BsaHI
CTX-M-5 (7), CTX-M-6 and CTX-M-7 (12), and Toho-1 (16).
and, in addition, showed the same restriction map for EcoRV,
TABLE 3. PCR-RFLP analysis to differentiate among genes highly related to bla
Length(s) (bp) of DNA fragment(s) obtained after digestion with:
500, 310, 90 a The percent identities between blaCTX-M-2 and Toho-1, CTX-M-4, CTX-M-5, CTX-M-6, and CTX-M-7 were 99.9, 98.1, 99.2, 97.5, and 98.4% respectively
(NALIGN program; PCGene; IntelliGenetics). PCR products from amplifications with PCTX-M-2 and whole DNA extracted from group A V. cholerae isolates and their
derivative transconjugants were incubated with restriction enzymes, and the fragment sizes (boldface type) were estimated after electrophoresis on a 1.8% agarose gel.
The sizes of the restriction fragments from the theoretical PCR products from amplifications with PCTX-M-2 and genes highly related to blaCTX-M-2 were deduced from
DISCUSSION
The most common mechanism of resistance to -lactam
antibiotics, including recent cephalosporins, is the productionof a -lactamase enzyme. Since 1971 transferable ampicillin
resistance have been described in V. cholerae isolates fromAfrica, Asia, and Latin America (10, 19, 32, 33, 42, 43, 44). However, only two plasmidic -lactamases have been identi-
fied to date. First, TEM-1 a broad-spectrum -lactamase was
identified in isolates from Asia and Africa (10, 33). Second,SAR-1, a carbenicillinase with ampicillin-hydrolyzing activitylower than that of TEM-1, was described in only two isolatesfrom Tanzania (33). In previous work (34), we reported for thefirst time on the isolation of an ESBL-producing V. choleraestrain (i.e., isolate M1502). In the present work, in addition toa TEM-1-type -lactamase, we have identified two extended-
spectrum plasmid-borne enzymes not related to TEM- orSHV-type enzymes in V. cholerae O1 isolates recovered duringseveral cholera seasons in Argentina. For group A isolates, the
-lactam susceptibility profiles of the transconjugant strains,
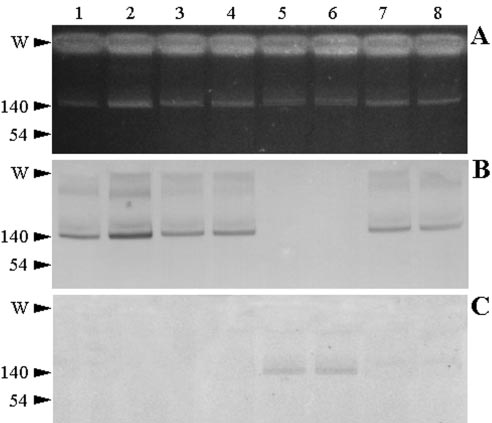
FIG. 1. Plasmid profiles and Southern blots of V. cholerae isolates
and their derivative transconjugants. Lanes: 1, 3, 5, and 7, V. cholerae
for which the cefotaxime/ceftazidime MIC ratio was 32, were
isolates M1502, M1516, M1573, and M3030, respectively; 2, 4, 6, and 8,
almost identical to those reported for strains producing several
E. coli transconjugants M1538, M3099, M1561, and M3033, respec-
CTX-M enzymes (16, 45), even though different E. coli recip-
tively. (A) Plasmid DNA was extracted as indicated in Materials and
ient strains (C600 and ER1793) were used in the mating ex-
Methods and analyzed on a 0.7% agarose gel. (B) The gel was trans-
ferred and blotted onto a nylon membrane and hybridized against a
periments. The results of IEF and PCR-RFLP assays suggest
900-bp PCR product from group A isolates as a probe. (C) The mem-
the presence of either CTX-M-2 or a highly related enzyme
brane was stripped and reprobed with the 740-bp PCR product from
different from those grouped in the CTX-M-2 cluster already
the group B isolate. The locations of the loading wells (W) and refer-
reported (5, 45). Otherwise, for the group B isolate, the -lac-
ence plasmids (numbers indicate sizes [in kilobases]) are shown on the
tam susceptibility profile of the transconjugant strain, for
which the cefotaxime/ceftazidime MIC ratio was 0.06, was verysimilar to those reported for strains producing PER enzymes
HincII, PstI, and SphI compared to that for the bla
(26; A. Bauernfeind et al., Abstr. 34th Intersci. Conf. Antimi-
crob. Agents Chemother., abstr. C74, 1994). These data, in
conjunction with the results of IEF and PCR-RFLP analysis,
PER primers generated amplification products (740 bp)
only with the templates from the group B V. cholerae isolate
are consistent with the presence of a PER-2-type enzyme in V.
and its transconjugant. Because of the high degree of DNA
cholerae M1561 and its transconjugant, M1573. Cefoxitin inhi-
bition of this enzyme, performed on the IEF gel before the
assay did not allow differentiation between such genes. A PCR-
iodometric method was carried out, allowed the identification
RFLP analysis carried out with both group B amplification
products and HindIII, PstI, and SphI showed a unique restric-
The expression of CTX-M-type and PER-2-type enzymes in
tion fragment profile which was identical to that expected for
V. cholerae isolates resulted in MICs of -lactams lower than
those commonly observed for CTX-M- or PER-2-producing
PER-2 gene and different from that expected for the
strains that belong to the family Enterobacteriaceae (with the
PER-1 gene (data not shown). The use of PTEM primers with
all DNA templates produced amplification products of 500 bp,
exception of the MICs for -lactams which are poor substrates
which is of the expected size of products from a gene encoding
for these enzymes, such as cefoxitin and imipenem). This fact
was more evident with ESCs, such as cefotaxime, ceftazidime,
and cefepime, for which the MICs were 32, 32, and 16 times
Plasmid profiles and locations of genes encoding ESBLs.
higher, respectively, for the E. coli transconjugants than for the
Analysis of the plasmid profiles of V. cholerae M1502, M1516,
V. cholerae isolates. Therefore, differences between V. cholerae
M1573, and M3030 and their respective transconjugants
and enterobacteria would exist in terms of the levels of expres-
(M1538, M3099, M1561, and M3033) showed the presence of
sion of ESBLs, the susceptibilities of penicillin-binding pro-
single plasmids of about 150 kb in each strain (Fig. 1A). The
teins to such -lactam agents, or the periplasmic concentra-
DNA hybridization analysis with the 900-bp PCR product from
group A isolates as a probe located the gene encoding the
Genetic determinants conferring resistance to non--lactam
CTX-M-type enzyme on the megaplasmids of group A isolates
antimicrobial agents were also encoded by the 150-kb plasmid
and their respective transconjugants (Fig. 1B). Reprobing of
harbored in each V. cholerae isolate. Multidrug resistance in V.
the same membrane with the 740-bp PCR product from the
cholerae O1 isolates has so far been linked to plasmids, and the
group B isolate as a probe showed that the megaplasmid of this
presence of self-transmissible multidrug-resistant plasmids
isolate and its transconjugant harbored the gene encoding the
from 100 to 200 kb has been reported in V. cholerae O1 isolates
from Asia, Africa, and Ecuador (10, 14, 19, 33, 40, 41, 43) and,
EXTENDED-SPECTRUM -LACTAMASES IN V. CHOLERAE
more recently, in isolates from Albania, Italy, and Brazil (11,
showed reduced susceptibility to nitrofurantoin, which was a
15). However, plasmids such those harbored by isolates M1516
common characteristic of isolates from the sixth cholera epi-
and M3030, which encode resistance to gentamicin, kanamy-
demic in Argentina, but was susceptible to all other antibiotics
cin, amikacin, streptomycin, trimethoprim-sulfamethoxazole,
tested. Furthermore, an E. coli strain isolated from the urine of
tetracycline, and chloramphenicol and, in the case of M1516,
the second case patient in the outbreak (the first one from
to erythromycin, in addition to -lactam antibiotic resistance,
whom an ESC-resistant V. cholerae strain had been isolated)
have not previously been reported. Moreover, to the best of
showed the same resistance phenotype as that for transconju-
our knowledge, gentamicin resistance was reported only in V.
gant strain M3033. These data suggest that multidrug-resistant
cholerae O1 isolates with two different resistance patterns (R
V. cholerae isolates could have resulted from the acquisition of
types) from two small outbreaks that occurred in Dacca, Ban-
the 150-kb plasmid encoding multidrug resistance in a conju-
gladesh, in 1981 (41) and in a single nosocomial pediatric ward
gation event with E. coli. The fact that CTX-M-2 is the most
in Samutsakorn, Thailand, in 1982 (40). In this work we have
frequent ESBL observed in E. coli and members of the family
reported on gentamicin resistance in V. cholerae O1 isolates
Enterobacteriaceae in Argentina (31, 35; Galas et al., 38th
with four different R types from two distantly occurring cholera
ICAAC; Galas et al., 39th ICAAC; Pasterán et al., 39th
seasons (4 years between the outbreaks). In addition, all the
ICAAC; A. C. Gales, T. M. Lewis, J. M. Casellas, V. Prado, J.
ESBL-producing V. cholerae isolates were coresistant to gen-
Smayevsky, and R. N. Jones, 39th Intersci. Conf. Antimicrob.
tamicin, as has been observed for ESBL-producing enterobac-
Agents Chemother., abstr. 1487, 1999) supports this hypothe-
terial isolates from Argentina (Galas et al., 38th ICAAC; Galas
sis. In addition, the risks for the potential spread of antimicro-
et al., 39th ICAAC; Pasterán et al., 39th ICAAC). This fact
bial resistance genes from V. cholerae to other bacteria have
suggests that both resistance determinants may be easily co-
been pointed out (19). Whatever the case, the findings of this
work point out the broad spread of ESBLs among genera
The MICs of tetracycline and chloramphenicol for both V.
belonging to families other than the Enterobacteriaceae. cholerae isolates resistant to such drugs (M1516 and M3030)
were 32- to 16-fold lower than those for their respective E. coliACKNOWLEDGMENTS
transconjugant strains (M3099 and M3033, respectively),
We thank Norma Binztein and Marta Rivas for providing epidemi-
whereas no differences in susceptibilities to aminoglycosides,
ologic data from the Argentinean cholera seasons; Marı´a Inés Caffer
trimethoprim, and sulfisoxazole were observed between the
for serotyping the V. cholerae isolates; and Marisa Rodrı´guez, Gustavo
original hosts and the derived transconjugants. This differential
López, and Ezequiel Tuduri Franco for technical assistance.
expression of tetracycline and chloramphenicol resistance
REFERENCES
mechanisms between V. cholerae and E. coli has been reported
1. Arlet, G., and A. Philippon. 1989. Construction by polymerase chain reaction
previously (30). This fact, in addition to the low levels of
and intragenic DNA probes for three main types of transferable -lactama-
resistance to -lactams discussed above, may be due to differ-
ses (TEM, SHV, CARB). FEMS Microbiol. Lett. 82:19–26.
2. Bauernfeind, A., I. Stemplinger, R. Jungwirth, S. Ernst, and J. M. Casellas.
ences between the E. coli envelope and that of V. cholerae,
1996. Sequences of -lactamase genes encoding CTX-M-1 (MEN-1) and
which could lack a permeability barrier or an active multidrug
CTX-M-2 and relationship of their amino acid sequences with those of other
efflux mechanism that enhances antibiotic input into the cell.
-lactamases. Antimicrob. Agents Chemother. 40:509–513.
3. Bauernfeind, A., I. Stemplinger, R. Jungwirth, P. Mangold, S. Amann, E.
Although replacement of the fluid lost in the stool remains
Akalin, Ö. Ang, C. Bal, and J. M. Casellas. 1996. Characterization of -lac-
the crucial element for the treatment of patients with cholera,
tamase gene blaPER-2, which encodes an extended-spectrum class A -lacta-
antimicrobial therapy is also important, since this can reduce
mase. Antimicrob. Agents Chemother. 40:616–620.
4. Bennish, M. L. 1994. Cholera: pathophysiology, clinical features and treat-
the volume of stool purged during illness, as well as shorten the
ment, p. 229–255. In I. K. Wachsmuth, P. A. Blake, and Ø. Olsvik (ed.),
duration of symptoms and the excretion of vibrios in the feces
Vibrio cholerae and cholera: molecular to global perspectives. American
Society for Microbiology, Washington, D.C.
(4). In addition, antimicrobial drugs are also used as chemo-
5. Bonnet, R., C. Dutour, J. L. M. Sampaio, C. Chanal, D. Sirot, R. Labia, C.
prophylaxis in close human contacts in attempts to control the
de Champs, and J. Sirot. 2001. Novel cefotaximase (CTX-M-16) with in-
epidemic spread from patients found to have cholera. To date,
creased catalytic efficiency due to substitution Asp-2403Gly. Antimicrob.
Agents Chemother. 45:2269–2275.
seven drugs have been recommended for the treatment of
6. Bonnet, R., J. L. Sampaio, R. Labia, C. de Champs, D. Sirot, C. Chanal, and
cholera: ampicillin, doxycycline (a long-acting form of tetracy-
J. Sirot. 2000. A novel CTX-M beta-lactamase (CTX-M-8) in cefotaxime-
cline), co-trimoxazole, erythromycin, furazolidone, chloram-
resistant Enterobacteriaceae isolated in Brazil. Antimicrob. Agents Che-
mother. 44:1936–1942.
phenicol, and norfloxacin (4). Multidrug-resistant plasmids
7. Bradford, P. A., Y. Yang, D. Sahm, I. Grope, D. Gardovska, and G. Storch.
harbored by isolate M1516 or M3030 carried genetic determi-
1998. CTX-M-5, a novel cefotaxime-hydrolyzing -lactamase from an out-
break of Salmonella typhimurium in Latvia. Antimicrob. Agents Chemother.
nants that conferred resistance to four (isolate M3030) or five
42:1980–1984.
(isolate M1516) of such first-line antimicrobial agents. The
8. Bush, K., G. A. Jacoby, and A. A. Medeiros. 1995. A functional classification
clinical relevance of these facts has not been established and
scheme for -lactamases and its correlation with molecular structure. Anti-
microb. Agents Chemother. 39:1211–1233.
deserves special attention when one is using prophylaxis mea-
9. de Champs, C., D. Sirot, C. Chanal, R. Bonnet, J. Sirot, and The French Study Group. 2000. A 1998 survey of extended-spectrum -lactamases in
In this work we have also reported on the first cholera
Enterobacteriaceae in France. Antimicrob. Agents Chemother. 44:3177–3179.
10. Dupont, M.-J., M. Jouvenot, G. Couetdic, and Y. Michel-Briand. 1985. De-
outbreak due to an ESBL-producing isolate. The outbreak
velopment of plasmid-mediated resistance in Vibrio cholerae during treat-
comprised 24 reported cases in a community of sugarcane
ment with trimethoprim-sulfamethoxazole. Antimicrob. Agents Chemother. 27:280–281.
workers in Irigoyen and Pichanal, Argentina, which are north
11. Falbo, V., A. Carattoli, F. Tosini, C. Pezzella, A. M. Dionisi, and I. Luzzi.
of Salta Province. Interestingly, the V. cholerae O1 strain iso-
1999. Antibiotic resistance conferred by a conjugative plasmid and a class I
lated from the first cholera patient in the outbreak (a 1-year-
integron in Vibrio cholerae O1 El Tor strains isolated in Albania and Italy.
Antimicrob. Agents Chemother. 43:693–696.
old male who acquired the illness during a hospitalization)
12. Gazouli, M., E. Tzelepi, A. Markogiannakis, N. J. Legakis, and L. S. Tzou- velekis. 1998. Two novel plasmid-mediated cefotaxime-hydrolyzing -lacta-
30. Ouellette, M., G. Gerbaud, and P. Courvalin. 1988. Genetic, biochemical
mases (CTX-M-5 and CTX-M-6) from Salmonella typhimurium. FEMS Mi-
and molecular characterization of strains of Vibrio cholerae multiresistant to
crobiol. Lett. 165:289–293.
antibiotics. Ann. Inst. Pasteur Microbiol. 139:105–113.
13. Gazouli, M., E. Tzelepi, S. V. Sidorenko, and L. S. Tzouvelekis. 1998. Se-
31. Power, P., M. Radice, C. Barberis, C. de Mier, M. Mollerach, M. Maltagli-
quence of the gene encoding a plasmid-mediated cefotaxime-hydrolyzing
atti, C. Vay, A. Famiglietti, and G. Gutkind. 1999. Cefotaxime-hydrolysing
class A -lactamase (CTX-M-4): involvement of serine 237 in cephalosporin
beta-lactamases in Morganella morganii. Eur. J. Clin. Microbiol. Infect. Dis.
hydrolysis. Antimicrob. Agents Chemother. 42:1259–1262. 18:743–747.
14. Glass, R. I., M. I. Huq, J. V. Lee, E. J. Threlfall, M. R. Khan, A. R. M. A.
32. Rahal, K., G. R. Gerbaud, and Y. A. Chabbert. 1973. Caractérisation d’un Alim, B. Rowe, and R. J. Gross. 1983. Plasmid-borne multiple drug resistance
facteur de résistance transférable de Vibrio cholerae biotype eltor. Ann.
in Vibrio cholerae serogroup O1, biotype El Tor: evidence for a point-source
Microbiol. (Inst. Pasteur) 124B:283–294.
outbreak in Bangladesh. J. Infect. Dis. 147:204–209.
33. Reid, A. J., and S. G. B. Amyes. 1986. Plasmid penicillin resistance in Vibrio
15. Hofer, E., B. R. Quintaes, E. M. dos Reis, D. dos P. Rodrigues, L. M. Seki, cholerae: identification of new -lactamase SAR-1. Antimicrob. Agents Che-
I. S. Feitosa, L. H. Ribeiro, and M. R. Ferreira. 1999. Emergencia da
mother. 30:245–247.
multipla resistencia a antimicrobianos em Vibrio cholerae isolados de paci-
34. Rossi, A., M. Galas, N. Binztein, M. Rivas, M. I. Caffer, A. Corso, M. Radice,
entes com gastroenterite no Ceara, Brasil. Rev. Soc. Bras. Med. Trop. 32: and G. Gutkind. 1993. Unusual multiresistant Vibrio cholerae O1 El Tor in
Argentina. Lancet 342:1172–1173.
16. Ishii, Y., A. Ohno, H. Taguchi, S. Imajo, M. Ishiguro, and H. Matsuzawa.
35. Rossi, A., H. Lopardo, M. Woloj, A. M. Picandet, M. Mariño, M. Galas, M.
1995. Cloning and sequence of the gene encoding a cefotaxime-hydrolyzing
Radice, and G. Gutkind. 1995. Non-typhoid Salmonella spp. resistant to
class A -lactamase isolated from Escherichia coli. Antimicrob. Agents Che-
cefotaxime. J. Antimicrob. Chemother. 36:697–702.
mother. 39:2269–2275.
36. Sabaté, M., R. Tarragó, F. Navarro, E. Miró, C. Vergés, J. Barbé, and G.
17. Jacoby, G. A., and A. A. Medeiros. 1991. More extended-spectrum -lacta- Prats. 2000. Cloning and sequence of the gene encoding a novel cefotaxime-
mases. Antimicrob. Agents Chemother. 35:1697–1704.
hydrolyzing beta-lactamase (CTX-M-9) from Escherichia coli in Spain. An-
18. Kariuki, S., J. E. Corkill, G. Revathi, R. Musoke, and C. A. Hart. 2001.
timicrob. Agents Chemother. 44:1970–1973.
Molecular characterization of a novel plasmid-encoded cefotaximase (CTX-
37. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: a
M-12) found in clinical Klebsiella pneumoniae isolates from Kenya. Antimi-
laboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, Cold
crob. Agents Chemother. 45:2141–2143.
19. Kruse, H., H. Sørum, F. C. Tenover, and Ø. Olsvik. 1995. A transferable
38. Simarro, E., F. Navarro, J. Ruiz, E. Miro, J. Gómez, and B. Mirelis. 2000.
multiple drug resistance plasmid from Vibrio cholerae O1. Microb. Drug
Salmonella enterica serovar Virchow with CTX-M-like -lactamase in Spain.
Resist. 1:203–210.
J. Clin. Microbiol. 38:4676–4678.
20. Labia, R., and M. Barthélémy. 1979. L’enzymogramme des beta-lactamases:
39. Southern, E. M. 1975. Detection of specific sequences among DNA frag-
adaptation en cel de la methode iodometrique. Ann. Microbiol. (Paris)
ments separated by gel electrophoresis. J. Mol. Biol. 98:503–517. 130B:295–304.
40. Tabtieng, R., S. Wattanasri, P. Echeverria, J. Seriwatana, L. Bodhidatta, A.
21. Medeiros, A. A. 1997. Evolution and dissemination of -lactamases acceler- Chatkaeomorakot, and B. Rowe,. 1989. An epidemic of Vibrio cholerae El
ated by generations of -lactam antibiotics. Clin. Infect. Dis. 24(Suppl.
Tor Inaba resistant to several antibiotics with a conjugative group C plasmid
1):19–45.
coding for type II dihydrofolate reductase in Thailand. Am. J. Trop. Med.
22. Nakamura, M., S. Sato, T. Ohya, S. Suzuki, and S. Ikeda. 1986. Plasmid
Hyg. 41:680–686.
profile analysis in epidemiological studies of animal Salmonella typhimurium
41. Threlfall, E. J., and B. Rowe. 1982. Vibrio cholerae El Tor acquires plasmid-
infection in Japan. J. Clin. Microbiol. 23:360–365.
encoded resistance to gentamicin. Lancet i:42.
23. National Committee for Clinical Laboratory Standards. 2000. Methods for
42. Threlfall, E. J., B. Rowe, and I. Huq. 1980. Plasmid-encoded multiple anti-
dilution antimicrobial susceptibility tests for bacteria that grow aerobically,
biotic resistance in Vibrio cholerae El Tor from Bangladesh. Lancet i:1247–
5th ed., vol. 20, no. 2. Approved standard M7-A5. National Committee for
Clinical Laboratory Standards, Wayne, Pa.
43. Threlfall, E. J., B. Said, B. Rowe, and A. Dávalos-Pérez. 1993. Emergence of
24. National Committee for Clinical Laboratory Standards. 2000. Performance
multiple drug resistance in Vibrio cholerae O1 El Tor from Ecuador. Lancet
standards for antimicrobial disk susceptibility tests, 7th ed., vol. 20, no. 1. 342:1173.
Approved standard M2-A7. National Committee for Clinical Laboratory
44. Towner, K. J., N. J. Pearson, and F. O’Grady. 1979. Resistant Vibrio cholerae
El Tor in Tanzania. Lancet ii:147–148.
25. Nordmann, P., and T. Naas. 1994. Sequence analysis of PER-1 extended-
45. Tzouvelekis, L. S., E. Tzelepi, P. T. Tassios, and N. J. Legakis. 2000. CTX-
spectrum -lactamase from Pseudomonas aeruginosa and comparison with
M-type -lactamases: an emerging group of extended-spectrum enzymes.
class A -lactamases. Antimicrob. Agents Chemother. 38:104–114.
Int. J. Antimicrob. Agents 14:137–142.
26. Nordmann, P., E. Ronco, T. Naas, C. Duport, Y. Michel-Briand, and R.
46. Vahaboglu, H., L. M. C. Hall, L. Mulazimoglu, S. Dodanli, I. Yildirim, and Labia. 1993. Characterization of a novel extended-spectrum -lactamase D. M. Livermore. 1995. Resistance to extended-spectrum cephalosporins,
from Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 37:962–969.
caused by PER-1 -lactamase, in Salmonella typhimurium from Istanbul,
27. Oliver, A., J. C. Pérez-Dı´az, T. M. Coque, F. Baquero, and R. Cantón. 2001.
Turkey. J. Med. Microbiol. 43:294–299.
Nucleotide sequence and characterization of a novel cefotaxime-hydrolyzing
47. Vahaboglu, H., R. Öztürk, G. Aygün, F. Coskunkan, A. Yaman, A. Kaygusuz,
-lactamase (CTX-M-10) isolated in Spain. Antimicrob. Agents Chemother. H. Leblebicioglu, I. Balik, K. Aydin, and M. Otkun. 1997. Widespread 45:616–620.
detection of PER-1 type extended-spectrum -lactamases among nosoco-
28. Olsvik, Ø., T. Popovic, and P. I. Fields. 1993. PCR detection of toxin genes
mial Acinetobacter and Pseudomonas aeruginosa isolates in Turkey: a nation-
in strains of Vibrio cholerae O1, p. 266–270. In D. H. Persing, T. F. Smith,
wide multicenter study. Antimicrob. Agents Chemother. 41:2265–2269.
F. C. Tenover, and T. J. White (ed.), Diagnostic molecular microbiology.
48. Yamamoto, T., G. B. Nair, M. J. Albert, C. C. Parodi, and Y. Takeda. 1995.
American Society for Microbiology, Washington, D.C.
Survey of in vitro susceptibilities of Vibrio cholerae O1 and O139 to antimi-
29. Olsvik, Ø., and N. A. Strockbine. 1993. PCR detection of heat-stable, heat-
crobial agents. Antimicrob. Agents Chemother. 39:241–244.
labile, and shiga-like toxin genes in Escherichia coli, p. 271–276. In D. H.
49. Yan, J. J., W. C. Ko, S. H. Tsai, H. M. Wu, Y. T. Jin, and J. J. Wu. 2000.
Persing, T. F. Smith, F. C. Tenover, and T. J. White (ed.), Diagnostic
Dissemination of CTX-M-3 and CMY-2 -lactamases among clinical isolates
molecular microbiology. American Society for Microbiology, Washington,
of Escherichia coli in southern Taiwan. Antimicrob. Agents Chemother. 38:4320–4325.
Yleistä n lasten kivunhoito kokenut valtaisan muutoksen viimeisen 15 vuoden aikana n vielä 1980- luvulta raportteja, joissa lapsia lääkittiin leikkauksen jälkeen vähän tai ei lainkaan n edellytykset kivun aistimiseen kehittyvät 24. –26. raskausviikkoon mennessä à kipua hoidettava n huonosti hoidettu kipu aiheuttaa lapselle pitkäkestosia, jopa elinikäisiä, käyttäytymisen / ps
FO GUANG SHAN NAN TIEN TEMPLE RISK ASSESSMENT FOR SCHOOL TOURS Potential risks Activity List hazards/risks related Control Strategies Please list to each activity/program Outline strategies for ensuring visitor safety for this potential risk and the venue 1. Guided 1.1,1.2 &1.3 Wear closed in shoes with non slip sole 1.1 Stairs,
 DISCUSSION
DISCUSSION